A few days back, sparc objected:
How often have we seen this very thread before? I am not interested in fishing but even I realize that I’ve seen the Abu 6500 C3 reel before (according to Google it appears 42 times on this site). Just opening another thread will not bring the stillborn FSCO/I to life. Didn’t you read what WE had to say about it? And what about Dembski, Meyer, Behe, Marks et al.? Do you think they even consider FSCO/I? FSCO/I just dead and never lived.
The substance of this is of course that I have repeatedly used an exploded view of the Abu 6500 C3 reel:
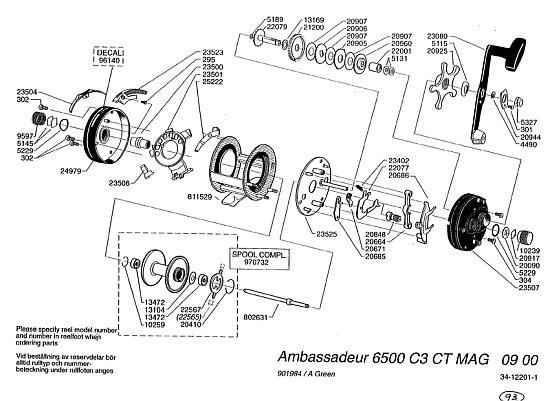
. . . as an apt, concrete example of the hard, undeniably factual reality of Wicken wiring diagram-based, functionally specific complex interactively organised entities and associated information (which is what the acronym FSCO/I represents: functionally specific, complex organisation and/or associated information — info that may be quantified by using a string of structured y/n q’s to describe the wiring diagram. Which, in the end is what AutoCAD etc do.)
FSCO/I is real, it aptly describes a feature of familiar reality.
The remaining question, is, is it relevant to the world of the living cell?
And again, I have provided repeatedly three cases in point that establish that:
first, a simplified piping and instrumentation diagram of an oil refinery (a classic wiring diagram):

second, the cell’s metabolic reaction network in outline:
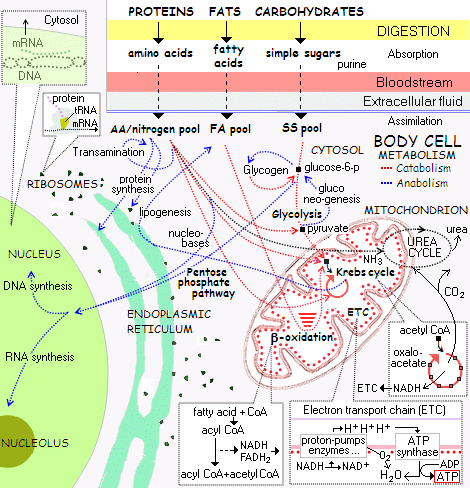
third, the top left corner blown up and filled in, the protein assembly process which is algorithmic, code driven and based on the information stored in DNA:
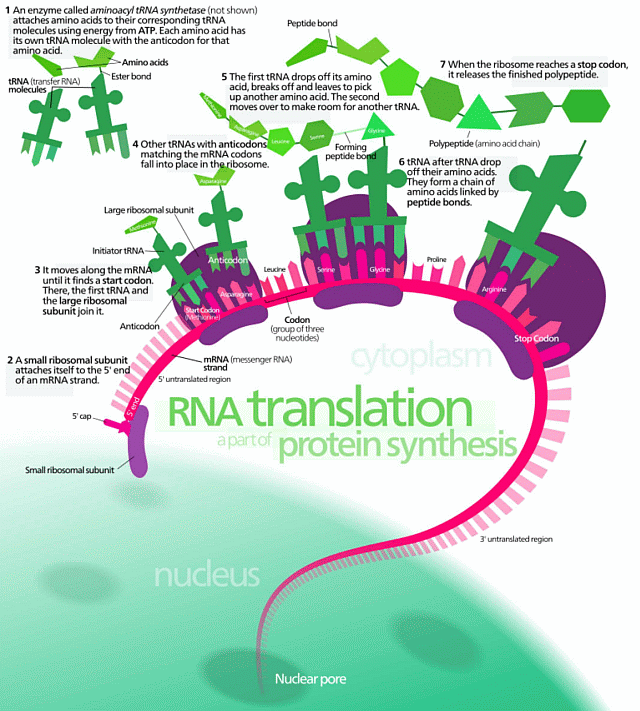
This is already enough to show that FSCO/I is relevant and real, at least to an unbiased person who is not troubled to the point of obvious distress by a further astonishing fact. Namely, that on a trillion-member observational basis, FSCO/I is reliably produced by intelligently directed configuration. Indeed, the only empirically observed adequate cause of FSCO/I is design.
So, we are epistemically entitled, on the simple logic of scientific induction, to infer that FSCO/I is a strong sign of design as cause.
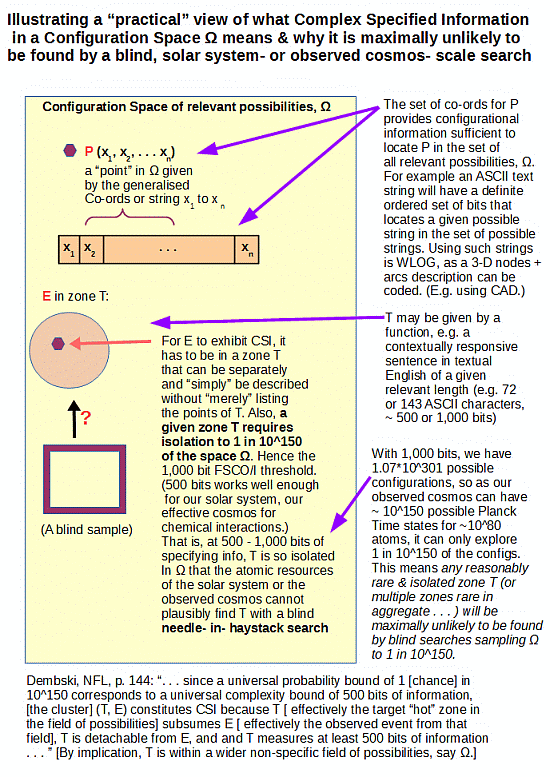
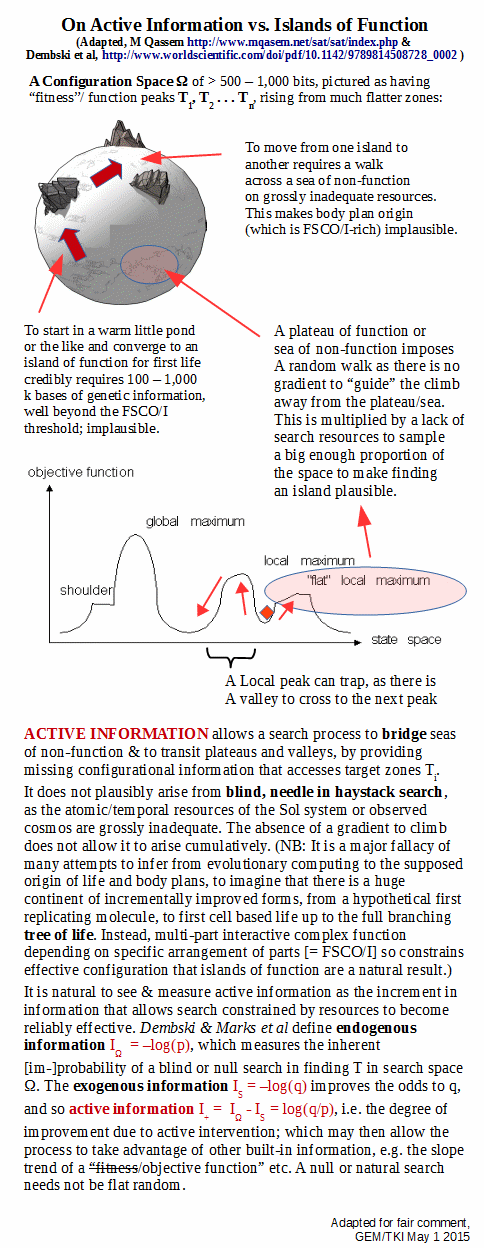
A point that is easily backed up by analysing the challenge posed by blind needle in haystack search, once we pass 500 – 1,000 bits of functionally specific complexity:
 Or, as I noted in the original post where the objection by sparc appears:
Or, as I noted in the original post where the objection by sparc appears:
. . . let me clip a summary of the needle-to-stack challenge:
Our observed cosmos has in it some 10^80 atoms, and a good atomic-level clock-tick is a fast chem rxn rate of perhaps 10^-14 s. 13.7 bn y ~10^17 s. The number of atom-scale events in that span in the observed cosmos is thus of order 10^111.
The number of configs for 1,000 coins (or, bits) is 2^1,000 ~ 1.07*10^301.
That is, if we were to give each atom of the observed cosmos a tray of 1,000 coins, and toss and observe then process 10^14 times per second, the resources of the observed cosmos would sample up to 1 in 10^190 of the set of possibilities.
It is reasonable to deem such a blind search, whether contiguous or a dust, as far too sparse to have any reasonable likelihood of finding any reasonably isolated “needles” in the haystack of possibilities. A rough calc suggests that the ratio is comparable to a single straw drawn from a cubical haystack ~ 2 * 10^45 LY across. (Our observed cosmos may be ~ 10^11 LY across, i.e. the imaginary haystack would swallow up our observed cosmos.)
Of course, as posts in this thread amply demonstrate the “miracle” of intelligently directed configuration allows us to routinely produce cases of functionally specific complex organisation and/or associated information well beyond such a threshold. For an ASCII text string 1,000 bits is about 143 characters, the length of a Twitter post.
As just genomes for OOL start out at 100 – 1,000 k bases and those for OOBP credibly run like 10 – 100+ mn bases, this is a toy illustration of the true magnitude of the problem.
So, FSCO/I is real, is relevant and on solid inductive grounds points to the best explanation of FSCO/I being, design. And if someone imagines that the fact that cells replicate themselves makes a crucial difference, let him or her ponder the fact that in his Nat Theol, 1804, Paley’s mention of stumbling across a watch in a field was just introductory. It is in Ch 2 that he went for his deeper argument. Let me clip, noting how he anticipated the idea of a von Neumann Kinematic self-replicator:
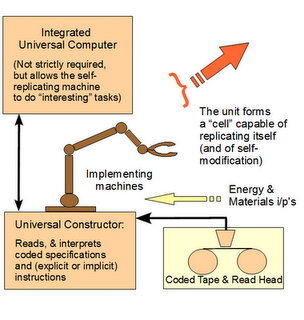
Suppose, in the next place, that the person who found the watch should after some time discover that, in addition to all the properties which he had hitherto observed in it, it possessed the unexpected property of producing in the course of its movement another watch like itself — the thing is conceivable; that it contained within it a mechanism, a system of parts — a mold, for instance, or a complex adjustment of lathes, baffles, and other tools — evidently and separately calculated for this purpose . . . .
In short, the additional FSCO/I involved in a self-replication facility — which must also be explained — simply underscores the force of the point. And of course, William Paley also addressed the underlying concept behind the abbreviation of a descriptive phrase, FSCO/I: the relation of [a complex, organised interactive array] of its parts to their use.
All of this would be so, even if I were the only person in the world to make such an argument.
Even, if it only ever appeared in a blog or two.
But in fact, part of sparc’s dismissive case is to suggest that the concept, facts and reasoning just noted, have escaped the attention of even leading design thinkers. So, it has to be dubious to the point that no one needs to pay it any attention.
Of course, this is already a serious fallacy, the negative appeal to blind modesty in the face of authority. To which, the direct answer is, while we must rely on authorities for 99% of our reasoning and knowledge base, no authority is better than his/her facts, assumptions and reasoning, so to the merits we must ever go.
However, the suggestion is materially false, which was pointed out in comment 148 in the same thread (not that it has made one whit of difference to sparc for coming on a hundred further comments — he is still happily pounding out his dismissive talking points, drumbeat style).
Accordingly, I replied at fair length, which I now headline for record:
________________
KF, 148: >> . . . How eager — suspiciously eager — you are to not look at what is in front of you: concrete cases in point illustrating the reality and characteristics of functionally specific complex organisation and associated information, FSCO/I; and of course, to write a stillbirth certificate.
Utterly revealing of the underlying problem.
For, this is a blatant case of knocking over a strawman and pretending the strawman was all that ever was there. Backed up, by a hoped for negative appeal to authority in the teeth of the actual demonstration of the reality of FSCO/I before your eyes that you want to avert your eyes and mind from, complaining that you have seen these cases before.
(That is, with all due respect, you inadvertently showed how your root problem is the fallacy of the closed, indoctrinated mind. A particularly virulent form of selective hyperskepticism that has led you to disbelieve the testimony of your eyes. As the just linked explains. [I add: those who would caricature that other phrase I have often used as a descriptive phrase for what Simon Greenleaf long ago termed the error of the skeptic, should note that it describes a real problem encountered live also: inconsistent, double standards of required warrant that project a demand on what one is inclined to reject that one would not accept for a comparable case one is inclined to accept. And, where typically, if the hyperskeptical standard were to be generally applied whole fields of learning or common sense useful or even vital knowledge would vanish.])
Since you are trying to suggest that functionally specific complex organisation and associated information, FSCO/I, is a non-issue . . . predictable, given a recent exchange in another thread alluded to above, I will AGAIN cite the remark made by Dembski in No Free Lunch, where he highlighted that the functional subset of complex specified information is the relevant one for the biological world. That is, the concept just described and abbreviated, FYI, does appear at a crucial point in the writings of Dr Dembski:
p. 148:“The great myth of contemporary evolutionary biology is that the information needed to explain complex biological structures can be purchased without intelligence. My aim throughout this book is to dispel that myth . . . . Eigen and his colleagues must have something else in mind besides information simpliciter when they describe the origin of information as the central problem of biology.
I submit that what they have in mind is specified complexity [[cf. here below], or what equivalently we have been calling in this Chapter Complex Specified information or CSI . . . .
Biological specification always refers to function. An organism is a functional system comprising many functional subsystems. . . . In virtue of their function [[a living organism’s subsystems] embody patterns that are objectively given and can be identified independently of the systems that embody them. Hence these systems are specified in the sense required by the complexity-specificity criterion . . . the specification can be cashed out in any number of ways [[through observing the requisites of functional organisation within the cell, or in organs and tissues or at the level of the organism as a whole. {Dembski cites:}
Wouters, p. 148: “globally in terms of the viability of whole organisms,”
Behe, p. 148: “minimal function of biochemical systems,”
Dawkins, pp. 148 – 9: “Complicated things have some quality, specifiable in advance, that is highly unlikely to have been acquired by ran-| dom chance alone. In the case of living things, the quality that is specified in advance is . . . the ability to propagate genes in reproduction.”
On p. 149, he roughly cites Orgel’s famous remark from 1973, which exactly cited reads:
In brief, living organisms are distinguished by their specified complexity. Crystals are usually taken as the prototypes of simple well-specified structures, because they consist of a very large number of identical molecules packed together in a uniform way. Lumps of granite or random mixtures of polymers are examples of structures that are complex but not specified. The crystals fail to qualify as living because they lack complexity; the mixtures of polymers fail to qualify because they lack specificity . . .
And, p. 149, he highlights Paul Davis in The Fifth Miracle: “Living organisms are mysterious not for their complexity per se, but for their tightly specified complexity.”] . . .”
p. 144: [[Specified complexity can be more formally defined:] “. . . since a universal probability bound of 1 [[chance] in 10^150 corresponds to a universal complexity bound of 500 bits of information, [[the cluster] (T, E) constitutes CSI because T [[ effectively the target hot zone in the field of possibilities] subsumes E [[ effectively the observed event from that field], T is detachable from E, and and T measures at least 500 bits of information . . . ”
Nor is this a novelty introduced by Dembski. When he would have been in High School, in 1973, FYFI, this is what leading OOL researcher, Leslie Orgel, had to say:
. . . In brief, living organisms [–> bio-functional context] are distinguished by their specified complexity. Crystals are usually taken as the prototypes of simple well-specified structures, because they consist of a very large number of identical molecules packed together in a uniform way. Lumps of granite or random mixtures of polymers are examples of structures that are complex but not specified. The crystals fail to qualify as living because they lack complexity; the mixtures of polymers fail to qualify because they lack specificity . . . .
[HT, Mung, fr. p. 190 & 196:] These vague idea can be made more precise by introducing the idea of information. Roughly speaking, the information content of a structure is the minimum number of instructions needed to specify the structure. [–> this is of course equivalent to the string of yes/no questions required to specify the relevant “wiring diagram” for the set of functional states, T, in the much larger space of possible clumped or scattered configurations, W, as Dembski would go on to define in NFL in 2002 . . . ] One can see intuitively that many instructions are needed to specify a complex structure. [–> so if the q’s to be answered are Y/N, the chain length is an information measure that indicates complexity in bits . . . ] On the other hand a simple repeating structure can be specified in rather few instructions. [–> do once and repeat over and over in a loop . . . ] Complex but random structures, by definition, need hardly be specified at all . . . . Paley was right to emphasize the need for special explanations of the existence of objects with high information content, for they cannot be formed in nonevolutionary, inorganic processes. [The Origins of Life (John Wiley, 1973), p. 189, p. 190, p. 196.]
And FYSYFI a few years later, J S Wicken in 1979, probably while Dembski was in College as an undergrad:
‘Organized’ systems are to be carefully distinguished from ‘ordered’ systems. Neither kind of system is ‘random,’ but whereas ordered systems are generated according to simple algorithms [[i.e. “simple” force laws acting on objects starting from arbitrary and common- place initial conditions] and therefore lack complexity, organized systems must be assembled element by element according to an [[originally . . . ] external ‘wiring diagram’ with a high information content . . . Organization, then, is functional complexity and carries information. It is non-random by design or by selection, rather than by the a priori necessity of crystallographic ‘order.’ [[“The Generation of Complexity in Evolution: A Thermodynamic and Information-Theoretical Discussion,” Journal of Theoretical Biology, 77 (April 1979): p. 353, of pp. 349-65. (Emphases and notes added. Nb: “originally” is added to highlight that for self-replicating systems, the blue print can be built-in.)]
The descriptive phrase, functionally specific complex organisation and associated information, FSCO/I for short, FYI, is directly based on Wicken’s remarks and is informed by a much wider circle of considerations.
You go on to call up Meyer, obviously having failed to note what he was already noted to have stated in answer to Falk’s hostile review of the 2009 book, Signature in the Cell:
For nearly sixty years origin-of-life researchers have attempted to use pre-biotic simulation experiments to find a plausible pathway by which life might have arisen from simpler non-living chemicals, thereby providing support for chemical evolutionary theory. While these experiments have occasionally yielded interesting insights about the conditions under which certain reactions will or won’t produce the various small molecule constituents of larger bio-macromolecules, they have shed no light on how the information in these larger macromolecules (particularly in DNA and RNA) could have arisen. Nor should this be surprising in light of what we have long known about the chemical structure of DNA and RNA. As I show in Signature in the Cell, the chemical structures of DNA and RNA allow them to store information precisely because chemical affinities between their smaller molecular subunits do not determine the specific arrangements of the bases in the DNA and RNA molecules. Instead, the same type of chemical bond (an N-glycosidic bond) forms between the backbone and each one of the four bases, allowing any one of the bases to attach at any site along the backbone, in turn allowing an innumerable variety of different sequences. This chemical indeterminacy is precisely what permits DNA and RNA to function as information carriers. It also dooms attempts to account for the origin of the information—the precise sequencing of the bases—in these molecules as the result of deterministic chemical interactions . . . .
[[W]e now have a wealth of experience showing that what I call specified or functional information (especially if encoded in digital form) does not arise from purely physical or chemical antecedents [[–> i.e. by blind, undirected forces of chance and necessity]. Indeed, the ribozyme engineering and pre-biotic simulation experiments that Professor Falk commends to my attention actually lend additional inductive support to this generalization. On the other hand, we do know of a cause—a type of cause—that has demonstrated the power to produce functionally-specified information. That cause is intelligence or conscious rational deliberation. As the pioneering information theorist Henry Quastler once observed, “the creation of information is habitually associated with conscious activity.” And, of course, he was right. Whenever we find information—whether embedded in a radio signal, carved in a stone monument, written in a book or etched on a magnetic disc—and we trace it back to its source, invariably we come to mind, not merely a material process. Thus, the discovery of functionally specified, digitally encoded information along the spine of DNA, provides compelling positive evidence of the activity of a prior designing intelligence. This conclusion is not based upon what we don’t know. It is based upon what we do know from our uniform experience about the cause and effect structure of the world—specifically, what we know about what does, and does not, have the power to produce large amounts of specified information . . .
When you can cogently address such, you will rise above strawman tactics.
But in fact, your strawman errors go beyond that.
For, the focal issue addressed in the OP is not “old hat.” It is specifically the role of active bridging information in the origin of FSCO/I, required to bridge seas of non-function to arrive at OOL and OOBP in the relevant configuration spaces. {Added, May 8: As in . . . }
Worse, this is a case of refusing to notice what is in front of you.
The 6500 fishing reel is emblematic of literally trillions of cases of FSCO/I all around you, including in the PC you used to compose your dismissive comment and to read this one in reply. FSCO/I is real.
The same phenomenon is exhibited in the digitally coded text s-t-r-i-n-g in your dismissive statement, which directly manifests the kind of specific functional organisation found in DNA, mRNA and onward in proteins and enzymes assembled through the FSCO/I rich process and systems in the ribosome. All, backed up by the further FSCO/I in the metabolic networks of the living cell.
All of which are illustrated in the OP for this thread, and all of which you are ever so eager to sweep away by resorting to strawman tactics . . . >>
________________
So, while obviously the acronym FSCO/I and the similar ones FSCI and dFSCI for the informational part and the digitally coded informational part are obviously my own (though GP often uses dFSCI here at UD and elsewhere), the concept has long been recognised and as shown it is real and relevant. Just as the concept of selective hyperskepticism is also real and relevant. Likewise, evolutionary materialism. All of which I have championed here at UD and elsewhere. Indeed, there is a “new” one too, OOBP — for, origin of body plans.
Now, I also took time to look up several other cases in the literature, which I will now clip as a chain of pearls:
KF, 185: >> Just as a fresh perspective on complex biological, coded functional information, we may wish to ponder:
http://www.ncbi.nlm.nih.gov/pm…..MC3319427/
Journal ListTheor Biol Med Modelv.9; 2012PMC3319427
Theor Biol Med Model. 2012; 9: 8.
Published online 2012 Mar 14. doi: 10.1186/1742-4682-9-8
PMCID: PMC3319427Dichotomy in the definition of prescriptive information suggests both prescribed data and prescribed algorithms: biosemiotics applications in genomic systems
David J D’Onofrio,corresponding author1 David L Abel,corresponding author2 and Donald E Johnson3
Bioinformatics has opened up the field of molecular biology through the use of computer science and statistics. Data mining of genetic information includes discovering relationships between individual DNA sequences and variability in disease [1]. More importantly, the application of computer science will contribute to identifying intricate complex data and algorithmic structures that are part of the biological processes that manage and maintain metabolic functions of the cell.
Biological organisms are considered to be controlled and regulated by Functional Information (FI) [2-8]. FI comes closer to expressing the intuitive and semantic sense of the word “information” than mere Shannon combinatorial uncertainty or reduced uncertainty (poorly termed “mutual entropy”). The innumerable attempts that have been made to reduce the functional information of genomics and molecular biology to nothing more than physical combinatorics and/or thermodynamics will fail for reasons best summarized in the peer-reviewed anthology entitled The First Gene: The Birth of Programming, Messaging and Formal Control [9].
“Functional Information (FI)” has now been formalized into two subsets: Descriptive Information (DI) [7] and Prescriptive Information (PI) [7,10,11]. This formalization of definitions precludes the prevailing confusion of informational terms in the literature. The more specific and accurate term “Prescriptive Information (PI)” has been championed by Abel [12-16] to define the sources and nature of programming controls, regulation and algorithmic processing. Such prescriptions are ubiquitously instantiated into all known living cells [13]. PI either instructs or produces formal function [12] in such a way as to organize and institute a prescribed set of logic-gate programming choices. Without such steering of physicochemical interactions by “Choice-Contingent Causation and Control” (CCCC) [17-19], metabolic pathways and cycles would be impossible to integrate into a cooperative and holistic metabolism. The Organization (O) Principle [19] states that nontrivial formal organization can only be produced by CCCC. [–> Cf OP]
Maynard Smith [20] argued that bioinformation is both specific and intentional. Maynard Smith also pointed out in this same paper the irreversibility of information transfer. Information moves only from signal to response, not in the reverse direction. He argued that genetic information implies the possibility of misinterpretation or error. Maynard Smith also considered genetic information to be undetermined by cause-and-effect necessity. But he considered genetic information to be gratuitous (not called for by the circumstances: unwarranted) [20].
Jablonka [21] argues that life is dependent upon semantic information, and that Shannon “information” is insufficient to explain life. She emphasizes, as does Adami [22], the importance of “aboutness.” Aboutness relates to meaning which in biology relates to biofunction.
Jablonka [21] also argues that semantic information can only exist with living or designed systems. “Only a living system can make a source into an informational input.” On page 588 Jablonka emphasizes the function of bioinformation. Thus the joint authors of this paper are not alone in our emphasis on the formal nature of life’s many control mechanisms.
A closer examination of Prescriptive Information (PI) has led to a dichotomy in its definition to differentiate between 1) what are prescribed data, and 2) what are prescribed algorithms. As the concepts of computer science are applied to the cell, it is necessary to deconstruct information structures to identify and differentiate data from algorithms. The DNA polynucleotide molecule consists of a linear sequence of nucleotides, each representing a biological placeholder of adenine (A), cytosine (C), thymine (T) and guanine (G). This quaternary system is analogous to the base two binary scheme native to computational systems. As such, the polynucleotide sequence represents the lowest level of coded information expressed as a form of machine code . . . >>
KF, 186: >> . . . some more of the same,
http://www.ncbi.nlm.nih.gov/pm…..MC4092032/
C R Biol. 2011 Jan; 334(1): 1–5.
Published online 2010 Dec 30. doi: 10.1016/j.crvi.2010.11.008
PMCID: PMC4092032
NIHMSID: NIHMS561755Thermodynamic perspectives on genetic instructions, the laws of biology, diseased states and human population control
J. T. Trevors corresponding author and M. H. Saier, Jr.
This article examines in a broad perspective entropy and some examples of its relationship to evolution, genetic instructions and how we view diseases. Many knowledge gaps abound, hence our understanding is still fragmented and incomplete. Living organisms are programmed by functional genetic instructions (FGI), through cellular communication pathways, to grow and reproduce by maintaining a variety of hemistable, ordered structures (low entropy). Living organisms are far from equilibrium with their surrounding environmental systems, which tends towards increasing disorder (increasing entropy). Organisms must free themselves from high entropy (high disorder) to maintain their cellular structures [–> i.e. cellular, functional organisation] for a period of time sufficient enough to allow reproduction and the resultant offspring to reach reproductive ages. This time interval varies for different species. Bacteria, for example need no sexual parents; dividing cells are nearly identical to the previous generation of cells, and can begin a new cell cycle without delay under appropriate conditions. By contrast, human infants require years of care before they can reproduce. Living organisms maintain order in spite of their changing surrounding environment, that decreases order according to the second law of thermodynamics. These events actually work together since living organisms create ordered biological structures by increasing local entropy. From a disease perspective, viruses and other disease agents interrupt the normal functioning of cells. The pressure for survival may result in mechanisms that allow organisms to resist attacks by viruses, other pathogens, destructive chemicals and physical agents such as radiation. However, when the attack is successful, the organism can be damaged until the cell, tissue, organ or entire organism is no longer functional and entropy increases. . . . .
Atoms are ancient relics of the hypothesized Big Bang (Matsuno, 2008) and can be used to construct life forms under the control of FGIs (functional genetic instructions). Living organisms are programmed by FGIs, which flow through a biochemical communication pathway involving DNA–> RNA–> proteins, to instruct cells how to assemble into living organisms. They are programmed to grow and reproduce by maintaining a variety of hemistable, ordered structures (low entropy state) (Schrodinger, 1944). They are far from equilibrium with their surrounding environment, which tends towards increasing disorder (Dolev & Elitzur, 1998). This is achieved by absorption of energy, from our thermonuclear sun, which provides the energy for the conversion of inanimate material into living organisms. This occurs on our planet with conditions commensurate with the maintenance of the life forms that comprise our singular biosphere system (Dolev & Elitzur, 1998; Gatenby & Frieden, 2007).
Researchers have devoted time and effort to defining and understanding the characteristics of life, from the atomic to the biospherical levels of organization (Penzlin, 2009; Schrodinger 1944) and in more recent years the possibility of synthetic single-celled life. Biology can therefore be viewed as the study of life (and death) at all levels of biological organization . . . Science relies on the fundamental laws of thermodynamics in addition to the knowledge that: (1) the cell is the basic unit of life; (2) life arises only from life; (3) a cell is the only living structure that can grow and divide (Trevors, 2004), and (4) functional genetic instructions flow along a cellular communication pathway to provide the instructions for the challenges from entropy, with reproduction as the normal outcome. Although natural selection prevents many individual organisms from reproducing, others must succeed if a species is to survive, even though all individuals within a species die, generally just not at the same time.
The presence of the pattern of thought captured in the acronym FSCO/I should be abundantly apparent. >>
KF, 187: >> . . . let us continue:
http://www.ncbi.nlm.nih.gov/pubmed/18762901/
Naturwissenschaften. 2009 Jan;96(1):1-23. doi: 10.1007/s00114-008-0422-8. Epub 2008 Sep 2.
The riddle of “life,” a biologist’s critical view.
Penzlin H1.
Author information
AbstractTo approach the question of what life is, we first have to state that life exists exclusively as the “being-alive” of discrete spatio-temporal entities. The simplest “unit” that can legitimately be considered to be alive is an intact prokaryotic cell as a whole. In this review, I discuss critically various aspects of the nature and singularity of living beings from the biologist’s point of view. In spite of the enormous richness of forms and performances in the biotic realm, there is a considerable uniformity in the chemical “machinery of life,” which powers all organisms. Life represents a dynamic state; it is performance of a system of singular kind: “life-as-action” approach. All “life-as-things” hypotheses are wrong from the beginning. Life is conditioned by certain substances but not defined by them. Living systems are endowed with a power to maintain their inherent functional order (organization) permanently against disruptive influences. The term organization inherently involves the aspect of functionality, the teleonomic, purposeful cooperation of structural and functional elements. Structures in turn require information for their specification, and information presupposes a source. This source is constituted in living systems by the nucleic acids. Organisms are unique in having a capacity to use, maintain, and replicate internal information, which yields the basis for their specific organization in its perpetuation. The existence of a genome is a necessary condition for life and one of the absolute differences between living and non-living matter. Organization includes both what makes life possible and what is determined by it. It is not something “implanted” into the living beings but has its origin and capacity for maintenance within the system itself. It is the essence of life. The property of being alive we can consider as an emergent property of cells that corresponds to a certain level of self-maintained complex order or organization. >>
KF, 188: >> Just for hammering the point home:
http://www.ncbi.nlm.nih.gov/pubmed/18202877/
Theory Biosci. 2004 Jun;123(1):3-15. doi: 10.1016/j.thbio.2004.03.001.
Evolution of cell division in bacteria.
Trevors JT1.
Author information
AbstractMolecular evolution in bacteria is examined with an emphasis on cell division. For a bacterial cell to assemble and then divide required an immense amount of integrated cell and molecular biology structures/functions to be present, such as a stable cellular structure, enzyme catalysis, minimal genome, septum formation at mid-cell and mechanisms to take up nutrients and produce and use energy, as well as store it. The first bacterial cell(s) capable of division must have had complex cell and molecular biology functions. At this stage of evolution, they would not have been primitive cells but would have reached a threshold in evolution where cell division occurred in a regulated manner. >>
KF, 193: >> Let’s try a sample chapter:
6 SYSTEMS BIOLOGY OF CELL ORGANIZATION
The first few chapters of this textbook laid the foundation for understanding cell structure and function. We learned that life depends on organic molecules, which form the building blocks for macromolecules such as proteins, nucleic acids, and carbohydrates. In addition, we considered cell organ-ization at a higher level. Cells contain complex structures such as membranes, chromosomes, ribosomes, and a cytoskeleton. Eukaryotic cells have organelles that provide specialized com-partments to carry out various cellular functions.
In short, complex specifically functional organisation of the cell based on interaction of correct, correctly arranged parts is a basic fact of life. Just as the OP describes. And just as the acronym FSCO/I describes . . . >>
In short the concept is indubitably real and relevant. It also appears in Dembski and in Meyer, and is rooted in Orgel and Wicken. Onwards, there are references to it in a significant quantity of professional literature.
I will continue to use it, the very fact of such desperate measures to object to it underscores why. END