2000: Ontogeny, Phylogeny and the Origin of Biological Information
Ontogeny, Phylogeny and the Origin of Biological Information
(2000)
by John A. Davison
Rivista di Biologia (Biology Forum), forthcoming
This paper is dedicated to the memory of six great scientists
William Bateson
Leo S. Berg
Robert Broom
Richard B. Goldschmidt
Pierre Grassé
Otto Schindewolf
No sadder proof can be given by a man of his own littleness
than disbelief in great men.
— Thomas Carlyle
Contents
1. Introduction
2. Epigenesis and preformation
3. Has evolution been guided?
4. Conclusion
References
Abstract and keywords
1. Introduction
In 1984 I presented the semi-meiotic hypothesis of organic evolution. This was followed by three other papers which exposed the complete failure of virtually every aspect of the Darwinian model (Davison 1984 1987 1993 1998). Incorporating the conclusions of Otto Schindewolf (1896-1971), Richard B. Goldschmidt (1878-1958), William Bateson (1861-1926), Robert Broom (1866-1951), Pierre Grassé (1895-1985) and Leo S. Berg (1876-1950), the semi-meiotic hypothesis can be summarized as follows. Macroevolution is largely finished. Sexual reproduction is incapable of supporting trans-specific (macroevolutionary) change. Accordingly, all significant change was produced presexually involving the first meiotic division. The essential feature of these changes was due not to micromutations in the genes themselves, but rather to the way in which those genes express their effects which is dependent upon their arrangement within the structure of the chromosome (position effect). The first meiotic division is ideally suited to this form of reproduction as it can preserve the original karyotype and at the same time produce structural novelties instantaneously in homozygous form (Davison 1998). Thus, this model provides a rational explanation for the absence of intermediates both in contemporary and in fossil species. Such a mechanism does not depend on the production of new information but rather on information already present in the genome i.e. preformed. This perspective remains in complete accord with the remarkable resemblances we find underlying disparate species, from the ubiquity of ATP to the universal 9 plus 2 nature of the structure of cilia and flagella wherever they are found in the living world. It should also be noted that Schindewolf (1993), Goldschmidt (1940), Berg (1969) and Grassé (1973) all subscribed to preadaptation (preformation) during evolutionary change.
These considerations bring me to the subject of this paper. First, what is the nature and source of the preformed information? Second, how was it made manifest? In considering these two questions
I came, as have others, to the realization that there is an intimate relationship between ontogeny and evolution (phylogeny).
The Russian ichthyologist and zoogeographer Leo S. Berg was perhaps the first to indicate that evolution and the development of the individual were closely related. In a clear demonstration of his
rejection of Darwinism, he wrote:
The laws of the organic world are the same whether we are
dealing with the development of an individual (ontogeny) or
that of a paleontological series (phylogeny). Neither in
the one nor in the other is there room for chance.
Nomogenesis (1969), page 134
Note that Berg insisted that chance plays no role in either process.
Whatever an organism becomes is determined entirely by the orderly
unfolding of events that take place during its embryonic development.
I accept Berg’s judgment and elaborate on it to see where that
perspective takes us with respect to our understanding of the
two as yet unresolved problems of ontogeny and phylogeny.
2. Epigenesis and preformation
These two terms originated from observations on the nature of the
developmental process. It is obvious that all of the information
necessary to produce a unique human being is contained (preformed)
in a single cell, the fertilized egg, a mere few microns in diameter.
On the other hand, the development of the individual is largely
epigenetic. I will deal first with epigenesis and then with
preformation. Epigenesis refers to the necessity for development
to proceed in a definite serial fashion. For example, neurulation
or the formation of the central nervous system cannot proceed until
gastrulation or the formation of the primitive digestive system is
complete. Following that, the optic cup can be formed. Only after
that can lens induction take place. These ontogenetic events have
interesting phylogenetic (evolutionary) parallels.
One of the most significant events in vertebrate evolution was the invention of bone. Since bone is significantly more dense than the cartilage it replaced, bony vertebrates would have been relegated to
the bottom of the sea if it were not for the early development of the swim bladder, an outgrowth of the primitive gut. This hydrostatic organ made it possible for the animal to maintain neutral buoyancy.
The swim bladder is homologous with the vertebrate lung which in turn made possible the invasion of the land and the evolution of the higher vertebrate taxa (amphibians, reptiles, birds and mammals).
Accordingly, the invention of bone and the swim bladder both served to preadapt subsequent vertebrates for life on land. It is significant that the Chondrichthyes (sharks skates, rays and chimeras) have remained cartilaginous and accordingly less specialized which may account for the ongoing successful presence of this very primitive group.
When something is lost during evolution it is rarely replaced. Examples are the loss of limbs in reptiles and amphibians, or digits in many tetrapod vertebrates. Returning to the swim bladder example,
the darters, tiny members of the perch family of fishes, have lost the swim bladder, a loss which allowed them to successfully invade swiftly
flowing freshwater streams. The Darwinian view would be that this
was an adaptation to the stream environment. An alternative view
would be that, having lost the swim bladder, the darters sought out
or perhaps simply stumbled into the stream environment where they
would have a clear advantage over buoyant competitors. Also the
Darwinian interpretation would suggest that the swim bladder was
gradually reduced, yet there is no evidence whatsoever for that
assumption. It would seem that many evolutionary specializations
occur instantly without intermediate forms. Thus, just as ontogeny
goes forward in epigenetic fashion, so has evolution. The two
processes also share irreversibility as a cardinal feature.
It is when we come to preformation that things really become
interesting. I now suggest that just as ontogeny and evolution share
epigenetic features, so also may they share the characteristics we
associate with preformation. In short, I propose that the information
for virtually all of evolution may have been present from very early
in the onset of that process. I realize that this idea may seem
ludicrous at first glance, yet it remains compatible with an enormous
number of otherwise enigmatic observations from comparative biology.
It also avoids postulating Lamarckian devices for which no evidence
has been forthcoming.
Before presenting this material it is necessary to grapple with the
question of the origin of the information itself. If, as I suggest,
the information was preformed, someone or something had to put it
there. I am not the first to confront this problem. I quote Pierre
Grassé:
According to Darwinian doctrine and Crick’s central dogma, DNA
is not only the depository and distributor of the information
but its sole creator. I do not believe this to be true.
(Grassé’s emphasis)
Evolution of Living Organisms
(1973), page 224
Oddly, Grassé does not pursue the problem further. I think we
have an obligation to do so. It is my understanding that information
does not arise de novo but must have a source. While that source has
yet to be identified, its existence is suggested by a host of examples
from both the plant and animal kingdoms.
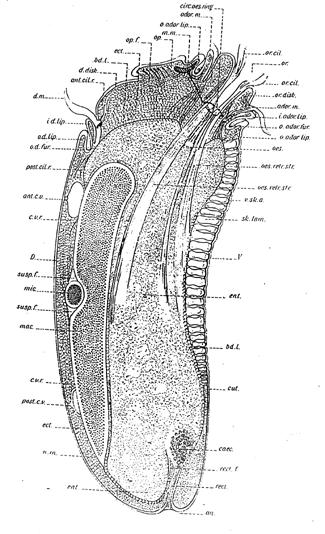
Figure 1. Diplodinium ecaudatum.
From Sharp (1914).
The unicellular Amoeba is structurally one of the simplest of the
Protozoa, yet in that same phylum can be found creatures exhibiting
strikingly advanced features. One such animal is Diplodinium
(Epidinium) ecaudatum(Figure 1). This ciliate exists in huge
numbers as symbionts in the stomachs of cattle (Sharp 1914). It has
a kind of “brain” (motorium), “circumoesophageal neural connectives”
resembling those of annelids and arthropods, clearly differentiated
“muscles”, a kind of “segmented skeleton” (skeletal laminae), a
“mouth”, “esophagus”, “rectum” and “anus” all elaborated within the
confines of a single cell. Such a creature not only demonstrates
that the information is already present for these structures, but
also casts considerable doubt on the notion that multicellularity
is a prerequisite for the division of labor. It is no wonder that
Libby Hyman described the protozoa as the acellular rather than the
unicellular animals. Why this creature should have such an array of
advanced features remains a complete mystery. Could it be there to
provide us with a clue to the nature of the evolutionary process?
The relict worm-like Peripatus with its curious combination of annelid
and arthropod characters could not be placed in either phylum, so it
has been placed in a phylum of its own, the Onycophora.
While we associate the placenta with the higher mammals, this
structure also is found in certain sharks. Berg (1969) discussed
this and related phenomena as examples of what he called physiological
acceleration. He noted that Peripatus also nourishes its embryos
with a kind of placenta. Thus Peripatus exhibits characters of three
taxa: the annelids, the arthropods and the placental mammals.
The primitive chordate Amphioxus (Branchiostoma lanceolatus), while
it has all three chordate features (dorsal hollow nervous system, gill
slits and notochord), has a kidney consisting of solenocytes of the
protonephridial (flame cell) type found in the Platyhelminthes, the
Aschelminthes and the polychaete Annelida. On the other hand, certain
oligochaete Annelida (earthworms) have a tubular kidney system more
like that of vertebrates. The limbless amphibia (Apoda) have large
yolk-laden eggs suspended by albuminous chalazae closely resembling
the situation in the cleidoic (shelled) eggs of reptiles and birds.
They lack only the shell. Such bizarre forms as the egg-laying
monotreme mammals (Platypus and Echidna) can suddenly seem acceptable
if one simply assumes that those combinations of characters were
available when those forms evolved. When it comes to the possible
combinations of features one can almost say — Anything goes!
While we associate imaginal discs with the complete metamorphosis of
the higher insects (Diptera, Hymenoptera, Homoptera and Lepidoptera),
imaginal discs are also involved in the metamorphosis of certain
marine worms (Nemertea), with the formation of the adult or imago
taking place within the body of the Pilidium larva which then splits
open to release the adult form much as does the pupa of a moth
or butterfly. Considerations like these and many others prompted
Leo Berg (1969) to conclude: “Evolution is in a great measure an
unfolding of preexisting rudiments.”
The same view was independently offered by Pierre Grassé (1973):
“The existence of internal factors affecting evolution has to be
accepted by any objective mind.”
The very word evolution derives from the Latin evolvo meaning
to unfold as the pages of a book. Needless to say, a book has,
by definition, already been written.
The plant kingdom abounds with examples in support of preformation.
Distantly related plant species often produce similar, if not
identical, chemical scents of anise, lemon, orange, pineapple,
cinnamon and a host of other flavorings so valuable to our cuisine.
There is even a chocolate scented member of the mint family!
The behavioral and structural reciprocities that often exist between
plants and their insect pollinators indicate that they may have been
sharing the same informational pool when those evolutionary events
took place. One of the most remarkable of such interactions was
described by Berg (1969). Certain stick and leaf insects (Phasmodea)
resemble the plants that they inhabit even to the extent that the eggs
of the insects closely resemble both internally and externally the
seeds of the plants (Umbelliferae) they inhabit and, like the seeds,
are scattered on the ground where they may remain for up to two years.
The tunic of the Urochordata (sea squirts) is composed of cellulose,
otherwise a polymer associated with the plant kingdom.
Rather than assuming independent inventions of these remarkable
parallels, it seems to me more reasonable to postulate these events
resulted from the activation (derepression) of an enormous yet clearly
limited stockpile of potentialities which were available when those
events took place.
These considerations cast a whole new perspective on what has been
called convergent evolution. The morphological similarities that
exist, especially in the skeletal systems, between placental wolves
and bears and their marsupial counterparts, to my mind, defy any
explanation based on the accidental evolution of these similarities
through chance events. Thus, what has been described as convergent
evolution is not that at all but the expression from preformed sources
of virtually identical morphologies. In this connection it is useful
to recall the words of R.C. Punnett:
Natural selection is a real factor in connection with mimicry,
but its function is to conserve and render preponderant an
already existing likeness, not to build up that likeness
through the accumulation of small variations, as is so
generally assumed.
(my emphasis)
Mimicry in Butterflies
(1915), page 152
One is compelled to ask — What is the source of the already existing
likeness?
How such transformations were effected remains, of course, a complete
mystery. Unfortunately, since macroevolution seems no longer to be
in progress, we may never be able to resolve that issue.
However intangible, the issue remains — What is the origin of the
preexisting information?
Questions such as this can probably never be resolved. Nevertheless,
one can still offer observations which relate to such metaphysical
considerations. Some of our greatest minds have been physicists.
Galileo, Newton, Pascal, Faraday and, in particular, Einstein all
have referred to God one way or another. Richard P. Feynman (1998)
has compared scientific discovery to a religious experience. I think
it is fair to say that we are still ignorant of the source of bright
ideas, insights and acts of creative genius.
It is interesting to compare our interpretation of the laws that
govern the inanimate world with those that may be operative in the
animate world. Leo Berg, as the complete title of his book indicates,
believed that all of both evolution and development was the result
of laws, a perspective with which I have (admittedly reluctantly)
been forced to agree. If not chance, it seems to me that the only
alternative is to agree with Berg. The existence of laws presumes
a law maker or makers. That in turn suggests purpose. Don’t
representatives in Congress have some purpose in mind when they
enact legislation?
No one denies the validity of Galileo’s equation which relates
the distance that a body falls to time, or Newton’s laws of motion,
or Einstein’s equation relating energy and mass. Why then must one
reject, as the Darwinians do, the suggestion that comparable laws
exist or have existed controlling the living world? Everyone
accepts gravitation and the equations associated with it, yet
no one yet understands the cause of gravity. Accordingly, neither
in religion nor in science does acceptance demand understanding.
Nevertheless, the Darwinians continue to insist that evolution is
the result only of chance events. Stephen J. Gould has recently
compared evolution to a drunk reeling back and forth between the
bar room wall and the gutter (Gould 1996 page 149). He has also
described intelligence as an evolutionary accident. I will only
comment that it was some accident!
Isn’t it conceivable that there may really be no conflict between
religion and evolutionary science? Isn’t it possible that both
might be correct? I have come to believe just that. Blaise Pascal
offered the following commentary as well as the solution:
Men despise religion. They hate it and are afraid it may
be true. The cure for this is first to show that religion is
not contrary to reason, but worthy of reverence and respect.
Finally, let me close with the following appropriate quotations from
my favorite intellectual, Albert Einstein:
I want to know God’s thoughts … the rest are details.
The most beautiful thing we can experience is the mysterious.
It is the source of all true Art and Science.
Science without religion is lame. Religion without science
is blind.
I shall never believe that God plays dice with the world.
I rest my case.
The greatest impediment to progress in the field of evolution resides
in the stubborn refusal of the Darwinians to entertain any possibility
that the living world might be subject to laws such as govern
virtually every aspect of the inanimate world. I find it fascinating
that it is the physicists who postulate God while biologists typically
remain atheists or agnostics. I offer the following explanation for
that curious situation. It is the physicists who have been able to
discover the several laws that serve to describe and thereby explain
their domain of the inanimate world. Thus they have been forced to
come to grips with the reality that laws must have an origin. By way
of contrast, most biological scientists have yet to acknowledge those
laws that so clearly operate in the living world, and so they reject
them as being unnatural or mystical and accordingly unacceptable.
I see no reason to make such distinctions. Laws are laws whether
or not we like them or understand them. Accordingly, I have found
the acceptance of the existence of preformed laws and information
to be a necessary and liberating asset in the ongoing search for
ultimate truth concerning the two greatest mysteries in all of
biological science: ontogeny and phylogeny. They are obviously
intimately related just as Leo Berg so clearly recognized long ago.
I now firmly believe that in solving the one we will solve the other.
BERG, L. (1969) Nomogenesis; or, Evolution Determined by Law.
M.I.T. Press, Cambridge. (Original Russian edition 1922.)
DAVISON, J.A. (1984) Semi-meiosis as an evolutionary mechanism.
J. Theor. Biol., 111: 725-735.
DAVISON, J.A. (1987) Semi-meiosis and evolution: a response.
J. Theor. Biol., 126: 379-381.
DAVISON, J.A. (1993) The blind alley: Its significance for
evolutionary theory. Rivista di Biologia (Biology Forum),
86: 101-110.
DAVISON, J.A. (1998) Evolution as a self-limiting process.
Rivista di Biologia (Biology Forum), 91: 199-220.
FEYNMAN, R.P. (1998) The Meaning of It All: Thoughts of a
Citizen Scientist. Addison-Wesley, Reading, Massachusetts.
GOLDSCHMIDT, R.B. (1940) The Material Basis of Evolution.
Yale University Press, New Haven.
GOULD, S.J. (1996) Full House: The Spread of Excellence from Plato
to Darwin. Harmony Books, New York.
GRASSÉ, P. (1977) Evolution of Living Organisms: Evidence
for a New Theory of Transformation. Academic Press, New York.
(Original French edition 1973.)
PUNNETT, R.C. (1915) Mimicry in Butterflies.
University Press, Cambridge.
SCHINDEWOLF, O. (1993) Basic questions in paleontology.
University of Chicago Press, Chicago. (Original German edition 1950.)
SHARP, R.G. (1914) Diplodinium ecaudatum, with an account of its
neuromotor apparatus. University of California Publications in
Zoology, 13: 43-123.
Accepting the evidence that evolution is largely finished and that
sexual reproduction is incapable of supporting macroevolution,
we deduce that macroevolutionary changes were produced presexually
through the cytological events associated with the first meiotic
division. This reproductive mode is ideally suited to the production
of new structural rearrangements of preexisting genetic information
instantaneously in homozygous form. These new arrangements (position
effects) produce new and discrete species. Thus, speciation results
not from new genetic information, but rather from information already
present (preformed). The several parallels that exist between
epigenesis and preformation in both ontogeny (development) and
phylogeny (evolution) are discussed. I propose that both of
these phenomena have proceeded through the selective activation
(derepression) of an enormous potential supply of information
already present at the onset of each of these biological phenomena.
Acceptance of these possibilities can serve to liberate us in our
quest for the ultimate truth concerning these two closely related
phenomena.
Keywords: phylogeny, ontogeny, evolution
(c) Copyright 2000 by John A. Davison.
Latest modification July 8, 2000.
This document and others by the same author may be found on-line at:
http://www.uvm.edu/~jdavison/