One of the advantages of having an academic background in philosophy is that you learn how to spot bad arguments. The origin of life is a subject where fallacious arguments proliferate. In this post, I’ve put together a list of a dozen common fallacies that bedevil scientific speculations regarding the origin of life. The next time you come across a paper on the origin of life in a science journal, you might like to check how many fallacies the author of the paper commits. For the sake of convenience, I’ve listed the fallacies here:
1. The fallacy of begging the question.
2. The fallacy of conflating the issues.
3. The fallacy of confusing the unknown with the undemonstrated.
4. The fallacy of large numbers.
5. The fallacy of invoking the infinite.
6. The fallacy of invoking a hidden intelligent designer.
7. The fallacy of the over-generalized description.
8. The fallacy of inferring possibility from picturability, a.k.a. the Pegasus fallacy.
9. The fallacy of ignoring the experimental evidence.
10. The fallacy of understating the problem.
11. The fallacy of adopting over-optimistic estimates.
12. The fallacy of arguing from unproven conjectures.
1. The fallacy of begging the question.
 |
Diagram of a cyanobacterium. An oft-cited 2008 paper by Dryden, Thomson and White assumes the existence of bacteria in order to demonstrate that their claim the entire suite of all possible functional proteins could have been tried out during the 4-billion-year history of life on Earth. But since bacteria (the simplest living things) require proteins in order to function in the first place, what the authors really need to explain is how Nature could have “hit upon” a functional protein before life arose. This is a particularly pressing problem for evolutionists, since as we’ll see below, only a tiny fraction (less than 1 in 1060) of all possible protein sequences are functional, and even 4 billion years wouldn’t have been enough time for a functional protein to have arisen by chance. Image courtesy of Kelvinsong and Wikipedia.
The first and most egregious fallacy regarding the origin of life is the fallacy of begging the question: since we’re here, life must have originated by some chemical process. This fallacy was committed by no less an authority than Professor John D. Sutherland, a chemist at the UK-based Medical Research Council’s Laboratory of Molecular Biology, the co-author of a 2009 article in Nature on the origin of life and the co-winner, along with former colleague Matthew Powner, of the 2012 Origin of Life Challenge. In an interview titled “Where did we come from?” (February 16, 2011) with scientist Neil deGrasse Tyson, host of the PBS show NOVASCienceNow, Professor Sutherland stated: “We’re here on the planet, and we must be here as a result of organic chemistry.” In other words, we’re here, so abiogenesis (the origin of life from non-living matter) must be possible somehow. Of course, that conclusion doesn’t follow unless one assumes all of the following premises:
(1) Life began at some point in time during the history of the universe;
(2) The complexity which characterizes life must have had some cause; and
(3) The universe is a causally closed system during the entire course of its history: nothing and no-one outside it can interact with it in any way.
Finally, in order to rule out intelligently guided organic chemistry explaining the origin of life via front-loading (as I presume Sutherland would wish to do), one would have to assume an additional premise:
(4) Neither the laws nor the initial conditions of our universe were set by an intelligent being.
The first premise can of course be ascertained scientifically, and the second premise is an admission that the complexity of life requires an explanation of some sort – a point which even New Atheists such as Richard Dawkins readily concede. However, the third and fourth premises can only be described as blatantly question-begging metaphysical assumptions. We don’t know that the universe is causally closed, and we don’t know that it was not designed. Professor Sutherland’s assertion that “we must be here as a result of organic chemistry” is therefore without warrant: it simply begs the question.
An even more egregious example of the fallacy of begging the question can be found in a 2008 paper by David Dryden, Andrew Thomson and John White of Edinburgh University, titled, How much of protein sequence space has been explored by life on Earth? (Journal of the Royal Society, Interface 6 August 2008, vol. 5 no. 25, pp. 953-956, doi: 10.1098/rsif.2008.0085), which defends the claim that during the course of life’s evolution, living organisms have tried out all possible functional proteins.
Two assumptions are generally made when considering the molecular evolution of functional proteins during the history of life on Earth. Firstly, the size of protein sequence space, i.e. the number of possible amino acid sequences, is astronomically large and, secondly, that only an infinitesimally small portion has been explored during the course of life on Earth…
We suggest that the vastness of protein sequence space is actually completely explorable during the populating of the Earth by life by considering upper and lower limits for the number of organisms, genome size, mutation rate and the number of functionally distinct classes of amino acids. We conclude that rather than life having explored only an infinitesimally small part of sequence space in the last 4 Gyr [4 billion years – VJT], it is instead quite plausible for all of functional protein sequence space to have been explored and that furthermore, at the molecular level, there is no role for contingency…
Hence, we hope that our calculation will also rule out any possible use of this big numbers ‘game’ to provide justification for postulating divine intervention (Bradley 2004; Dembski 2004).
I should point out in passing that neither of the authors (Bradley and Dembski) cited in the final sentence of Dryden, Thomson and White’s paper argued for “divine intervention”; all they argued for was the occurrence of Intelligent Design in Nature.
However, since Dryden, Thomson and White are attempting to obviate the need for an Intelligent Designer, it is not enough for them to show that “for all practical (i.e. functional and structural) purposes, protein sequence space has been fully explored during the course of evolution of life on Earth” (italics mine). But since even bacteria (the simplest living things) require proteins in order to function in the first place, what the authors really need to explain is how Nature could have “hit upon” a functional protein before life arose. As we’ll see below, it turns out that only a tiny fraction (less than 1 in 1060) of all possible protein sequences are functional. This fraction is so small that even 4 billion years wouldn’t have been enough time for a functional protein to have arisen by chance on the primordial Earth. In their attempt to rule out any requirement for “divine intervention,” Dryden, Thomson and White are putting the cart before the horse.
Dr. Cornelius Hunter, a graduate of the University of Illinois with a Ph.D. in Biophysics and Computational Biology, has also pointed out this fallacy in Dryden, Thomson and White’s paper, in a recent post over at his blog, Darwin’s God, titled, Response to Comments: Natural Selection Doesn’t Help, Gradualism is Out, and so is Evolution (July 2, 2011):
To defend their second claim, that evolution can easily search the entire protein sequence space, the evolutionists present upper and lower bound estimates of the number of different sequences evolution can explore.
Their upper bound estimate of 10^43 (a one followed by 43 zeros) is ridiculous. It assumes a four billion year time frame with 10^30 bacteria constantly testing out new proteins. First, even for an upper bound estimate their time frame is about two to three orders of magnitude too large. And furthermore, from where did these bacteria come? Bacteria need thousands of, yes, proteins. You can’t use bacteria to explain how proteins first evolved when the bacteria themselves require an army of proteins.
The lower bound of 10^21 is hardly any more realistic. The evolutionists continue to use the four billion year time frame. And they also continue to rely on the pre existence of an earth filled with a billion species of bacteria (with their many thousands of pre existing proteins).
Since its publication, Dryden, Thomson and White’s article has been cited by no less than 21 scientific papers. Remarkably, none of the papers’ authors seems to have noticed the fallacy in the article they cited.
2. The fallacy of conflating the issues.
 |
 |
Left: A schematic of the type III secretion-system needle-complex, a protein appendage found in several Gram-negative bacteria. It is composed of approximately 30 different proteins. Image courtesy of Pixie and Wikpedia.
Right: Evolutionary biologist Lynn Margulis (1938-2011), photographed at a conference in La Coruña, Spain, on November, 9, 2005. Margulis once remarked: “To go from a bacterium to people is less of a step than to go from a mixture of amino acids to a bacterium.” (The End of Science, by John Horgan. Addison-Wesley Publishing Company, Inc., 1996, pp. 140-141.) Image courtesy of Javier Pedreira and Wikipedia.
The second fallacy committed by biologists when discussing the origin of life is the fallacy of conflating the issues – in this case, the issues of abiogenesis and the evolution of life (whether as a result of natural selection, as Darwinists believe, or a mutation-driven process, as Professor Masatoshi Nei contends, is irrelevant here).
In an interview on the PBS show “Faith and Reason,” Professor Richard Dawkins argued that the origin of life was a relatively simple problem, as it was the subsequent process of evolution which performed most of the work of generating the complexity of life:
The origin of life — the key process in the origin of life was the arising of a self-replicating molecule. This was a very simple thing compared with what it’s given rise to. By far the majority of the work in producing the elegant complexity of life is done after the origin of life, during the process of evolution. There does remain the very first step — I don’t think it’s necessarily a bigger step than several of the subsequent steps, but it is a step. And it’s a step which we don’t fully understand — mainly because it happened such a long time ago, and under conditions when the Earth was very different. And so it’s not necessarily possible to simulate again the chemical conditions of the origin of life. There are various theories for how it might have happened. None of them is yet fully convincing. It may be that none of them ever will be, because it may be that we shall never know fully what the conditions were. But I don’t find it at all a deeply mysterious step.
However, there are sound logical arguments against conflating the problem of life’s origin with the problem of how the first living things could have evolved into complex organisms. Even if the scientific case for complex life-forms arising through an unguided process of evolution were absolutely airtight, it would prove absolutely nothing about the probability of abiogenesis. Demonstrating that life evolved by an unguided process does not prove that it originated by an unguided process.
In his interview with PBS, Professor Dawkins downplays the difficulty of the origin of life on Earth: it involved the generation of “a self-replicating molecule,” which was “a very simple thing,” compared to “the work in producing the elegant complexity of life.” Dawkins might have done well to heed a remark made by evolutionary biologist Lynn Margulis (1938-2011), who was interviewed by John Horgan in The End of Science (Addison-Wesley Publishing Company, Inc., 1996, pp. 140-141), that “To go from a bacterium to people is less of a step than to go from a mixture of amino acids to a bacterium.”
3. The fallacy of confusing the unknown with the undemonstrated.
 |
Charles Darwin in 1881. A copy made by John Collier (1850-1934) in 1883 of his 1881 portrait of Darwin. Given to the National Portrait Gallery, London in 1896. Image courtesy of the National Portrait Gallery and Wikipedia.
The third fallacy is the fallacy of confusing the unknown with the undemonstrated. It is often argued that even if we have no idea how life on Earth originated, this shouldn’t alarm us: after all, physicists had no explanation for how gravity worked until Einstein came up with his General Theory of Relativity in 1915. Charles Darwin himself was guilty of this fallacy when he wrote:
It is no valid objection that science as yet throws no light on the far higher problem of the essence or origin of life. Who can explain what is the essence of the attraction of gravity? No one now objects to following out the results consequent on this unknown element of attraction; notwithstanding that Leibnitz formerly accused Newton of introducing “occult qualities and miracles into philosophy.” (The Origin of Species by means of natural selection, or the preservation of favoured races in the struggle for life. London: John Murray. 6th edition, 1872. Chapter XV, page 421.)
The mechanism of gravity was entirely unknown in Darwin’s day, as was the alleged mechanism for the origin of life from non-living matter. Nevertheless, there is a vital difference between the two. The existence of gravity can be publicly demonstrated, whereas abiogenesis cannot. Gravity is scientifically measurable, but we have no evidence that life ever originated from non-living matter.
4. The fallacy of large numbers.
 |
An exceptionally clear picture of the stars in the night sky, taken by astronomer Håkon Dahle at ESO’s La Silla Observatory in the Atacama desert, Chile. Many people argue that the emergence of life somewhere in the cosmos is inevitable, given the vast number of stars in the universe. Image courtesy of Håkon Dahle and Wikipedia.
The fourth fallacy on my list is what I will call the fallacy of large numbers – also known as the fallacy of appealing to “deep time” and/or the vastness of the universe, in order to explain the origin of life. One hears this argument with boring regularity, in popular discussions of the probability of intelligent life in outer space: “Look at all the stars out there! They’ve got planets, and there appears to be nothing special about our planet: scientists have already found some earth-like planets in our neighborhood. Are you seriously going to tell me we’re the only planet in the entire cosmos that has intelligent life?” And if this argument holds true for intelligent life, it must also hold, a fortiori, for life itself.
A milder version of this fallacy can be found in the writings of scientists who contend that the origin of life in our universe on at least one occasion – on Earth, billions of years ago – is not all that surprising, given the vast size and age of the cosmos. Nobel Prize winner and Harvard University Professor George Wald committed this fallacy in a now-famous article titled, “The origin of life” (Scientific American 191(2):44–53, August 1954), where he wrote:
When we consider the spontaneous origin of a living organism, this is not an event that need happen again and again. It is perhaps enough for it to happen once. The probability with which we: are concerned is of a special kind; it is the probability that an event occur at least once. To this type of probability a fundamentally important thing happens as one increases the number of trials. However improbable the event in a single trial, it becomes increasingly probable as the trials are multiplied. Eventually the event becomes virtually inevitable…
Time is in fact the hero of the plot. The time with which we have to deal is of the order of two billion years. What we regard as impossible on the basis of human experience is meaningless here. Given so much time, the “impossible” becomes possible, the possible probable, and the probable virtually certain. One has only to wait: time itself performs the miracles…
The important point is that since the origin of life belongs in the category of at-least-once phenomena, time is on its side. However improbable we regard this event, or any of the steps which it involves, given enough time it will almost certainly happen at least once. And for life as we know it, with its capacity for growth and reproduction, once may be enough.
I should mention in passing that Professor Wald, in subsequent correspondence, added that he had “no strong personal prejudice against invoking God’s intervention in the origin of life,” pointing out that “[t]he Jesuit priest, John Turberville Needham, a great champion of spontaneous generation, believed that God created matter initially with the potentiality of spontaneously generating life.”
In a subsequent article in the Proceedings of the National Academy of Sciences, titled, “The Origins of Life” (PNAS, Aug 1964; 52(2): 595–611), Wald went further, and estimated that “The number of planets suitable for life in the already observed universe may be of the order of 1017” (p. 601), adding: “This number is so vast – even if it were reduced a million times – as to make it difficult to avoid the conclusion that life is widespread in the universe.”
The fallacy implicit in this conclusion is that it assumes that the probability of life’s originating on Earth exceeds some finite threshold. In Professor Wald’s 1964 article, the threshold he assumed was a probability of one in a million. If that were the case, then indeed, all we would have to do is wait for life to appear on suitably hospitable planets. The problem, however, is that in the absence of solid experimental data, we have no grounds for assuming that the probability of life originating from non-living matter exceeds one in a million. Professor Wald was simply engaging in idle speculation.
5. The fallacy of invoking the infinite.
 |
Giordano Bruno claimed that there were an infinite number of worlds. This derivative modern illustration of Giordano Bruno is taken from a modern version of “Livre du recteur” (1578), University of Geneva, which is thought to be based on an original drawing – in other words, this is a copy of a copy. Image courtesy of Wikipedia.
The fifth fallacy, which might be considered a “fallback position” for people who embrace the previous fallacy, is the fallacy of invoking the infinite – in other words, appealing to the infinite multiverse in order to explain the origin of life. In a peer-reviewed article, “The Cosmological Model of Eternal Inflation and the Transition from Chance to Biological Evolution in the History of Life” (Biology Direct 2 (2007): 15, doi:10.1186/1745-6150-2-15), evolutionary biologist Dr. Eugene Koonin claims that the emergence of even a basic replication-translation system on the primordial Earth is such an astronomically unlikely event that we would need to postulate a vast number of universes, in which all possible scenarios are played out, in order to make its emergence likely. Thus Koonin is forced to appeal to the multiverse, in order to render the origin of life probable. The scientific argument presented in Koonin’s peer-reviewed paper has been republished (in virtually identical form) in his recent book, The Logic of Chance: The Nature and Origin of Biological Evolution (Upper Saddle River: FT Press, 2011, ISBN 978-0-13-262317-9).
However, the multiverse hypothesis is plagued by two problems of its own: first, it merely shifts the fine-tuning problem up one level, as a multiverse capable of generating any life-supporting universes at all would still need to be fine-tuned; and second, even the multiverse hypothesis implies that a sizable proportion of universes (including perhaps our own) were intelligently designed. In an influential essay entitled, The Teleological Argument: An Exploration of the Fine-Tuning of the Universe (in The Blackwell Companion to Natural Theology, edited by William Lane Craig and J. P. Moreland, 2009, Blackwell Publishing Ltd.), Dr. Robin Collins offers a scientific explanation of why even a “multiverse-generator” would still fail to eliminate the need for fine-tuning:
In sum, even if an inflationary-superstring multiverse generator exists, it must have just the right combination of laws and fields for the production of life-permitting universes: if one of the components were missing or different, such as Einstein’s equation or the Pauli Exclusion Principle, it is unlikely that any life-permitting universes could be produced. Consequently, at most, this highly speculative scenario would explain the fine-tuning of the constants of physics, but at the cost of postulating additional fine-tuning of the laws of nature.
Additionally, physicist Paul Davies has argued that the multiverse hypothesis is just as “theological” as the hypothesis that there is a God, since it implies the existence of intelligently designed universes:
Among the myriad universes similar to ours will be some in which technological civilizations advance to the point of being able to simulate consciousness. Eventually, entire virtual worlds will be created inside computers, their conscious inhabitants unaware that they are the simulated products of somebody else’s technology. For every original world, there will be a stupendous number of available virtual worlds – some of which would even include machines simulating virtual worlds of their own, and so on ad infinitum.
Taking the multiverse theory at face value, therefore, means accepting that virtual worlds are more numerous than “real” ones. There is no reason to expect our world – the one in which you are reading this right now – to be real as opposed to a simulation. And the simulated inhabitants of a virtual world stand in the same relationship to the simulating system as human beings stand in relation to the traditional Creator.
Far from doing away with a transcendent Creator, the multiverse theory actually injects that very concept at almost every level of its logical structure. Gods and worlds, creators and creatures, lie embedded in each other, forming an infinite regress in unbounded space.
— Paul Davies, A Brief History of the Multiverse, New York Times, 12 April 2003.
6. The fallacy of invoking a hidden intelligent designer.
 |
The “successes” reported in origin-of-life experiments are invariably the result of intelligent chemists controlling each step of the process. Consequently, they prove absolutely nothing regarding the possibility of life originating via an unguided process. In this photo, synthetic chemist Julie Perkins works to link two molecules, each of which binds to two protein binding sites, to make a new molecule that will bind more strongly and securely to a specific toxin protein than the individual molecules can. Photo courtesy of Lawrence Livermore National Laboratory and Wikipedia.
The sixth fallacy, which is often overlooked by scientists when they are zealously promoting their preferred model of the origin of life, is the fallacy of invoking a hidden intelligent designer. If you are going to put forward a model of how life could have arisen from non-living matter via an unguided process, then you really cannot expect to be taken seriously if the model you propose assumes the existence of a designer.
I referred above to the article in Nature (May 14, 2009; 459(7244):239-42, doi: 10.1038/nature08013) by Matthew W. Powner, Beatrice Gerland & John D. Sutherland, titled, “Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions,” in which the authors proposed that nucleotides could have joined together to form from natural chemicals, and thus create life.
In 2009, I contacted Professor John Walton, a chemist and professor at the University of St Andrews, for his take on the experiment by Powner, Gerland and Sutherland, and received the following reply:
I have seen previous papers by Sutherland and co-workers on supposed prebiotically plausible syntheses. The Nature paper you mention is of much the same kind. It’s nice chemistry, carried out with the best reagents and lab equipment, supervised by intelligent chemists. They use pure components, separate and purify products using the most up to date methods. They control the pH, temperature, reaction medium, reaction mixture, contact time, and energy input. Although they claim their route is plausible under prebiotic conditions, nothing in their paper makes this believable. They haven’t solved the stereoselectivity problem. They haven’t solved the problems of chemoselectivity in assembly of the pyrimidine ribonucleotides and they don’t address the problem of the information needed to assemble the nucledotides in the right order. What they show is that with intelligent chemists supervising each step of the process, nucleotides can be accessed from some simple molecules. It is good evidence of intelligent design at work.
Dr. Stephen Meyer, the acclaimed author of Signature in the Cell, has also pointed out that the experiment by Powner, Garland and Sutherland relied on intelligent design, in a letter to the Times Literary Supplement, which its editors chose not to publish:
Starting with several simple chemical compounds, Powner and colleagues successfully synthesized a pyrimidine ribonucleotide, one of the building blocks of the RNA molecule.
Nevertheless, this work does nothing to address the much more acute problem of explaining how the nucleotide bases in DNA or RNA acquired their specific information-rich arrangements, which is the central topic of my book [Signature in the Cell: DNA and the Evidence for Intelligent Design]. In effect, the Powner study helps explain the origin of the “letters” in the genetic text, but not their specific arrangement into functional “words” or “sentences.”
Moreover, Powner and colleagues only partially addressed the problem of generating the constituent building blocks of RNA under plausible pre-biotic conditions. The problem, ironically, is their own skillful intervention. To ensure a biologically-relevant outcome, they had to intervene — repeatedly and intelligently — in their experiment: first, by selecting only the right-handed isomers of sugar that life requires; second, by purifying their reaction products at each step to prevent interfering cross-reactions; and third, by following a very precise procedure in which they carefully selected the reagents and choreographed the order in which they were introduced into the reaction series.
Thus, not only does this study not address the problem of getting nucleotide bases to arrange themselves into functionally specified sequences, but the extent to which it does succeed in producing biologically relevant chemical constituents of RNA actually illustrates the indispensable role of intelligence in generating such chemistry.
Sadly, the above-cited paper by Powner, Gerland and Sutherland is all too typical of scientific papers dealing with the origin of life, in assuming the occurrence of a highly artificial sequence of steps that needs to be set up by an intelligent being in the first place, in order to generate the molecules leading to the first life-forms.
7. The fallacy of the over-generalized description.
 |
A plasma lamp. Some researchers have seriously proposed that plasma-based life forms exist on other planets in the cosmos, and that they may even have existed on the primordial Earth. Image courtesy of Luc Viatour and Wikipedia.
The seventh fallacy that recurs in arguments surrounding the origin of life is the fallacy of the over-generalized description. Proponents of this fallacy argue that since life can elsewhere in the universe may be exotic and completely different to life on Earth, the probability of the emergence of life may be much, much higher than we imagine. “Life on other planets may be very different from life on Earth,” they say. “It may not contain DNA, like terrestrial life. It may not even be carbon-based. It may be utterly unlike anything that we can imagine. All it needs to be able to do is metabolize, grow and replicate. There are lots of things in Nature that might be able to do that. So the probability of finding life of some sort somewhere in the cosmos is likely to be quite high.”
This kind of thinking is exemplified in an article by David Cohen in New Scientist (17 September 2003), titled, “Plasma blobs hint at new form of life“, which suggested the possibility of a plasma-based form of life elsewhere in the cosmos, and even proposed that it may have preceded DNA-based life on Earth:
Physicists have created blobs of gaseous plasma that can grow, replicate and communicate – fulfilling most of the traditional requirements for biological cells. Without inherited material they cannot be described as alive, but the researchers believe these curious spheres may offer a radical new explanation for how life began.
Most biologists think living cells arose out of a complex and lengthy evolution of chemicals that took millions of years, beginning with simple molecules through amino acids, primitive proteins and finally forming an organised structure. But if Mircea Sanduloviciu and his colleagues at Cuza University in Romania are right, the theory may have to be completely revised. They say cell-like self-organisation can occur in a few microseconds…
Sanduloviciu grew spheres from a few micrometres up to three centimetres in diameter… Each sphere had a boundary made up of two layers – an outer layer of negatively charged electrons and an inner layer of positively charged ions…
A distinct boundary layer that confines and separates an object from its environment is one of the four main criteria generally used to define living cells. Sanduloviciu decided to find out if his cells met the other criteria: the ability to replicate, to communicate information, and to metabolise and grow.
He found that the spheres could replicate by splitting into two. Under the right conditions they also got bigger, taking up neutral argon atoms and splitting them into ions and electrons to replenish their boundary layers.
Finally, they could communicate information by emitting electromagnetic energy, making the atoms within other spheres vibrate at a particular frequency. The spheres are not the only self-organising systems to meet all of these requirements. But they are the first gaseous “cells”.
Sanduloviciu even thinks they could have been the first cells on Earth, arising within electric storms. “The emergence of such spheres seems likely to be a prerequisite for biochemical evolution,” he says…
…[P]erhaps the most intriguing implications of Sanduloviciu’s work are for life on other planets. “The cell-like spheres we describe could be at the origin of other forms of life we have not yet considered,” he says. Which means our search for extraterrestrial life may need a drastic re-think. There could be life out there, but not as we know it.
The problem with this line of thinking is that even if the possibility of exotic life-forms is real, the question of how carbon-based life-forms arose, and in particular, how DNA-based (or for that matter, RNA-based) life-forms arose, remains just as intractable as it was before. Exotic life-forms don’t help us to solve this problem.
At this point, proponents of exotic life might turn around and object that I am being unreasonable and asking for too much: they could argue that while the origin of life in general during the history of the cosmos is highly probable, the origin of any particular biological molecule (such as DNA) may nevertheless be vastly improbable, just as the probability of getting the particular hand of cards that you are dealt in a game of Snap is astronomically low. But this objection can be met if we can show that life on Earth satisfies a general description which makes no mention of any particular molecule. A scientist can then reasonably ask how life satisfying that general description could have possibly originated, especially if she has good grounds for believing that the probability of that kind of life originating via unguided chemical processes is vanishingly low.
And indeed, it is not difficult to find such a general description, which applies to all life-forms on Earth but makes no mention of any specific molecule. All life on Earth requires the existence of proteins, which perform a vast array of functions within living organisms. What’s more, living things require not only simple proteins but also large, complex proteins – and to make matters worse, there are no known gradual pathways to forming such proteins. The term “protein” is a general one: it applies to any long chain of amino acid residues, bonded together by peptide bonds, which can fold up and perform a useful function. The question of how large, complex proteins arose is a scientifically legitimate one, and its difficulty would in no way be diminished by the discovery of exotic, plasma-based life-forms.
8. The fallacy of inferring possibility from picturability, a.k.a. the Pegasus fallacy.
 |
Statue of Pegasus on the roof of Poznań Opera House. Some eminent origin-of-life researchers are guilty of committing what I call the “Pegasus fallacy”: the fallacy of assuming that something is possible simply because it is imaginable. We can all imagine a flying horse, but it is only when we ask ourselves, “How would it fly?” that we realize that such a horse is not really possible. Image courtesy of Radomil and Wikipedia.
The eighth fallacy that one repeatedly encounters in discussions of the origins of life is the fallacy of inferring possibility from picturability. I have elsewhere referred to this as the Pegasus fallacy: if something is imaginable, then it is possible. (We can all imagine Pegasus in our heads, but this mental picture is merely superficial, as it ignores the fundamental question of how a horse would fly in the first place. When we ask ourselves this question, we realize that the existence of Pegasus is impossible.) This fallacy of inferring possibility from picturability can be regarded as the converse of another fallacy, which Richard Dawkins refers to as the argument from incredulity, where someone declares an event to be impossible, simply because he cannot imagine how it could have happened. On this point, Dawkins is correct: our inability to mentally picture how an event might have happened tells us nothing about the possibility of its occurring in the real world. But by the same token, the mere fact that we can picture how an event might have happened tells us nothing about whether the actual occurrence of that event is a realistic possibility (i.e. one that exceeds some threshold of probability that would make its occurrence scientifically credible) or not. To answer that question, we need two things: a picture with the requisite level of detail, and some solid mathematics.
An example of this kind of thinking can be found in a PBS interview for NOVA online (May 3, 2004; posted July 1, 2004), in which Joe McMaster, producer of “Origins: How Life Began,” interviewed paleontologist Andrew Knoll, a professor of biology at Harvard and author of Life on a Young Planet: The First Three Billion Years of Life. During the course of the interview, Professor Knoll stated:
I can imagine that there was a time before there was life on Earth, and then clearly there was a time X-hundred thousand years or a million years later when there were things that we would all recognize as biological. But there’s no question that we must have gone through some intermediate stage where, had you been there watching them, you might have placed your bets either way.
So I can imagine that on a primordial Earth you would have replicating molecules — not particularly lifelike in our definition, but they’re really getting the machinery going. Then some of them start interacting together and pretty soon you have something a little more lifelike, and then it incorporates maybe another piece of nucleic acid from somewhere else, and by the accumulation of these disparate strands of information and activity, something that you and I would look at and agree “that’s biological” would have emerged.
The problem with this kind of airy speculation is that the picture Professor Knoll sketches is entirely lacking in the requisite chemical details, and has no mathematical calculations to demonstrate the feasibility of the chemical steps involved. When a scientist says “I can imagine,” the typical response of his or her peers will be, “Show me.” Later on in the interview, Professor Knoll acknowledged this problem himself:
…[I]t’s fairly easy to make simple sugars, molecules called bases which are at the heart of DNA, molecules called amino acids which are at the heart of proteins. It’s fairly easy to make some of the fatty substances that make the coverings of cells. Making all of those building blocks individually seems to be pretty reasonable, pretty plausible.
The hard part, and the part that I think nobody has quite figured out yet, is how you get them working together. How do you go from some warm, little pond on a primordial Earth that has amino acids, sugars, fatty acids just sort of floating around in the environment to something in which nucleic acids are actually directing proteins to make the membranes of the cell?
Somehow you have to get all of the different constituents working together and have basically the information to make that system work in one set of molecules, which then directs the formation of a second set of molecules, which synthesizes a third set of molecules, all in a way that feeds back to making more of the first set of molecules. So you end up getting this cycle. I’m not sure we’ve gotten very far down the road to understanding how that really happens.
Indeed.
9. The fallacy of ignoring the experimental evidence.
 |
Scientists in a laboratory of the University of La Rioja. A number of recent papers on the origin of life have relied on what evolutionary biologist John Maynard Smith (1920-2004) has disparagingly referred to as “fact-free science” – i.e. science without any experimental data to back it up. Image courtesy of Urcomunicacion and Wikipedia.
The ninth fallacy plaguing origin-of-life discussions is the fallacy of ignoring the experimental evidence. In an article titled, From Complexity to Perplexity, author John Horgan takes aim at the hypothesis developed by Stuart Kauffman, of the Santa Fe Institute, that artificial life can shed light on the origin of life on Earth:
…[E]volutionary biologist John Maynard Smith of the University of Sussex…, who pioneered the use of mathematics in biology, took an early interest in work at the Santa Fe Institute and has twice spent a week visiting there. But he has concluded that artificial life is “basically a fact-free science.” During his last visit, he recalls, “the only time a fact was mentioned was when I mentioned it, and that was considered to be in rather bad taste.”
Another example of this kind of thinking can be found in a paper by Martin A. Nowak and Hisashi Ohtsuki, titled, Prevolutionary dynamics and the origin of evolution (PNAS 2008 : 0806714105v1-0; doi: 10.1073/pnas.0806714105; published online September 12, 2008). The authors handily summarize their thesis as follows:
Traditionally, one thinks of natural selection as choosing between different replicators. Natural selection arises if one type reproduces faster than another type, thereby changing the relative abundances of these two types in the population. Natural selection can lead to competitive exclusion or coexistence. In the present theory, however, we encounter natural selection before replication. Different information carriers compete for resources and thereby gain different abundances in the population. Natural selection occurs within prelife and between life and prelife. In our theory, natural selection is not a consequence of replication, but instead natural selection leads to replication. There is “selection for replication” is replicating sequences have a higher abundance than nonreplicating sequences of a similar length. We observe that prelife selection is blunt: typically small differences in growth rates result in small differences in abundance. Replication sharpens selection: Small differences in replication rates can lead to large differences in abundance.
Commenting on the article in a post over at Evolution News and Views (August 22, 2011), Jonathan McLatchie hit the nail on the head in a pithy observation he made:
The model proposed in this paper is highly theoretical and speculative — with no substantive practical experimental research to back it up.
In his post, McLatchie highlighted an additional flaw of the article by Novak and Ohtsuki: its inadequate treatment of the biological concept of functional information. The authors’ failure to address a key concept in biology renders their mathematical calculations (which, to be fair, are quite rigorous) largely irrelevant:
It’s not entirely clear in what sense the authors are using the term “information,” nor that they understand it. They tell us that “prelife is a generative system that can produce information.” We are also told that “Evolution needs a generative system that can produce unlimited information. Evolution needs populations of information carriers.” They also tell us on the first page that they “can define a prebiotic chemistry that can produce any binary string and thereby generate, in principle, unlimited information and diversity.” Since when was a set of random strings of characters a sound definition of “information” — at least in any meaningful sense as applied to biology?
10. The fallacy of understating the problem.
 |
The protein hexokinase, with the much smaller molecules of ATP and the simplest sugar, glucose, shown in the top right corner for comparison. At present, evolutionists are still struggling to explain how relatively small molecules like ATP might have been generated on the primordial Earth. They are nowhere near solving the problem of how proteins formed, let alone how the first life arose. Image courtesy of Tim Vickers and Wikipedia.
A tenth fallacy which comes up again and again in scientific discussions of the origins of life is the fallacy of understating the problem. Five years ago, a paper by Matthew Powner, Beatrice Gerland and John Sutherland, titled, Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions (Nature 459:239-242), grabbed headlines around the world when one of the papers authors, Professor John D. Sutherland, announced that they had show how nucleotides, the building blocks of RNA, could have spontaneously assembled themselves in the conditions of the primitive earth. “Chemist Shows How RNA Can Be the Starting Point for Life,” screamed the New York Times, in an article by Nicolas Wade (May 13, 2009). A pertinent question at this point might be: how big is a nucleotide, anyway? And how big is the smallest known independent living organism? (I won’t count viruses here, as they are inert on their own, and can only replicate inside other organisms.)
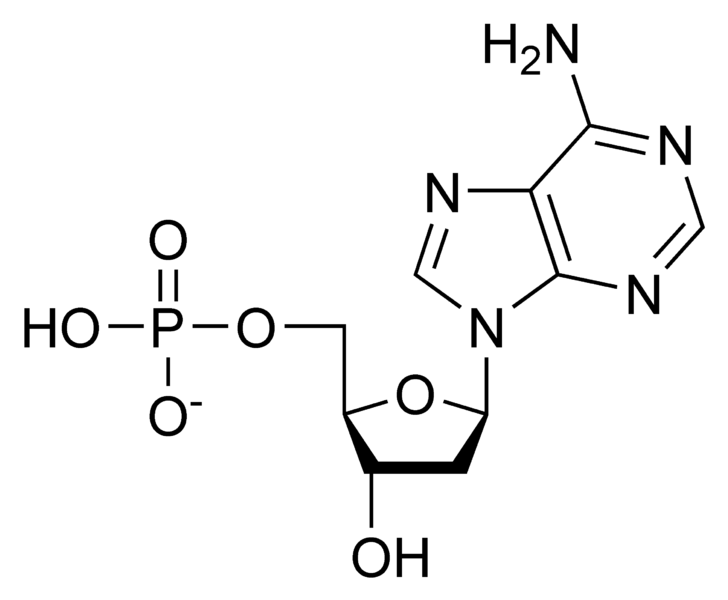 |
Chemical structure of a typical nucleotide, deoxyadenosine monophosphate. In 2009, Matthew Powner, Beatrice Gerland and John Sutherland announced that they had succeeded in synthesizing a nucleotide in the laboratory under conditions they claimed could have existed on the primordial Earth. However, a typical nucleotide has only a few dozen atoms, whereas the simplest known independent life-form contains at least one million times more atoms. Image courtesy of cacycle and Wikipedia.
The smallest known parasitic bacterium, M. genitalium has a molecular weight of 360,110 kilodaltons (kDa), or 360,110,000 daltons, while the smallest known free-living bacterium, Pelagibacter ubique, is more than twice as massive. (One dalton can roughly be defined as the mass of a typical hydrogen atom.) Printed in 10 point font, the letters of the M. genitalium JCVI-1.0 genome span 147 pages. By contrast, the RNA nucleotide which Sutherland et al. manufactured (ribo-cytidine phosphate) has a molecular weight of less than 360 daltons: less than a millionth that of the simplest bacterium.
The problem with the hype surrounding Sutherland’s paper should by now be readily apparent: it was totally out of proportion to the minor achievement that they had performed in the lab. The molecule they synthesized contained a mere two dozen or so atoms. By contrast, the number of atoms in an E. coli bacterium is around 7,000,000,000 – quite some difference!
Another example of the fallacy of understating the problem can be found in the ongoing scientific debate over how proteins might have arisen on the primordial Earth. Some experts have proposed that the first proteins may have been relatively short, making their emergence by random processes far more likely.
Dr. Cornelius G. Hunter is a graduate of the University of Illinois where he earned a Ph.D. in Biophysics and Computational Biology. In a recent personal communication, Dr. Hunter highlighted the problem with this proposal:
“Proteins are by no means created equal. They occupy a wide spectrum of size and complexity… Nor is there reason to think that evolution could live with the shorter, simpler ones at first, and then later somehow the larger, more complex ones would evolve. The larger ones appear to be needed, and there are not obvious gradual pathways to forming them… We’re still not close to the more complex proteins.”
11. The fallacy of adopting over-optimistic estimates.
The eleventh fallacy, closely related to the tenth, is the fallacy of adopting over-optimistic estimates.
Scientists now agree that only a very small proportion of all possible protein sequences has the property of being able to fold up and create a stable 3D structure. This finding severely undermines the claim that proteins could have arisen by an unguided process, as Dr. Ann Gauger explains in a post titled Protein Evolution: A Guide for the Perplexed:
…[T]he problem is that the number of possible protein sequences that could exist is very large, occupying a very large potential sequence space, but the number of proteins that do exist is much smaller, and they are widely scattered across sequence space (perhaps — in fact, that is one of the things being debated). The potential space is so large that a purely random search for rare functional proteins would spectacularly fail. So unless functional sequences are easy to find (very common), and/or are clustered together (easily reachable from one functional island to another), explaining current protein diversity without design is impossible.
Some origin-of-life researchers have attempted to argue that although the probability of finding a functional protein sequence by chance is very low, it is not astronomically low, and that there is sufficient time during the Earth’s history for such a sequence to have arisen naturally, without the need for any intelligent guidance. From an actual library of 6×1012 proteins each containing 80 contiguous random amino acids, Keefe and Szostak managed to isolate four ATP binding proteins and concluded that the proportion of all possible protein sequences that are actually functional might be as high as 1 in 1011, or 1 in 100,000,000,000. They concluded that functional proteins could therefore have arisen by an unguided, stochastic (i.e. random) process:
In conclusion, we suggest that functional proteins are sufficiently common in protein sequence space (roughly 1 in 1011) that they may be discovered by entirely stochastic means, such as presumably operated when proteins were first used by living organisms.
(Keefe AD, Szostak JW (2001). “Functional proteins from a random-sequence library.” Nature 410:715-718.)
Another team of scientists estimated that a random protein library of about 1024 members would be sufficient for finding one chorismate mutase molecule, making the problem of unguided natural processes hitting upon a functional sequence difficult but by no means impossible (Taylor SV, Walter KU, Kast P, Hilvert D (2001). “Searching sequence space for protein
catalysts.” Proceedings of the National Academy of Sciences USA 98:10596-10601. doi:10.1073/pnas.191159298.)
In a recent personal communication, Dr. Cornelius Hunter explained why these estimates of the probability of finding a functional protein sequence by chance are wildly over-optimistic, and pointed out that a more realistic estimate would be 1 in 1060, or 1 in 1 million million million million million million million million million million:
“First, Keefe and Szostak is not relevant as they were not seeking functional proteins, but merely mild ATP binding. Second, Taylor, et. al. deals with a simple, helix only, protein (homodimeric AroQ), biased the sequence toward helix forming amino acids and sequence patterns, did not fully randomize the sequence but only randomized regions, and is vague about how they arrive at their 10^24 tries required. Even if their calculation of 10^24 is reasonable, you’re dealing with a pretty simple protein… AroQ is toward the simple end of the spectrum… And finally there are several studies on slightly more complex, challenging proteins, all of which come in at around 10^60 – 10^80 attempts required.”
And as Dr. Hunter has pointed out (see fallacy #10 above), there are no good scientific grounds for thinking that life-forms based on shorter, simpler proteins could have existed in the beginning, giving rise to more advanced life-forms requiring larger, more complex ones later on. As he puts it: “The larger ones appear to be needed, and there are not obvious gradual pathways to forming them.”
If the more realistic, experimentally based estimate quoted by Dr. Hunter, that only 1 in 1060 to 1 in 1080 possible protein sequences are functional, is correct, then this would render the possibility of Nature hitting upon a functional protein via an unguided process astronomically unlikely – a point recognized by Taylor et al. in their paper:
Even a library with the mass of the Earth itself — 5.98 × 1027 g — would comprise at most 3.3 × 1047 different sequences, or a miniscule fraction of such diversity.
12. The fallacy of arguing from unproven conjectures.
 |
Proteins before and after folding. Image courtesy of Wikipedia.
The twelfth fallacy is the fallacy of arguing from unproven conjectures, and in so doing, imagining that one is making what scientists refer to as a bold prediction.
In their paper titled, How much of protein sequence space has been explored by life on Earth?, authors David Dryden, Andrew Thomson and John White (of Edinburgh University) defend the claim that “a reduced alphabet of amino acids is quite capable of producing all protein folds (approx. a few thousand discrete folds; Denton 2008) and providing a scaffold capable of supporting all protein functions… Therefore it is entirely feasible that for all practical (i.e. functional and structural) purposes, protein sequence space has been fully explored during the course of evolution of life on Earth.”
However, Dr. Cornelius Hunter has rebutted the arguments in Dryden, Thomson and White’s paper, in two posts over at his blog, Darwin’s God: Response to Comments: Natural Selection Doesn’t Help, Gradualism is Out, and so is Evolution (July 2, 2011) and The Amyloid Threat, Big Numbers Game and Quote Mining: Protein Evolution and How Evolutionists Respond to the Empirical Evidence (September 15, 2011). The key points from Dr. Hunter’s response are as follows:
The paper [by Dryden, Thomson and White] attempts to make two general points. First that evolution can succeed with a much smaller protein sequence space and second, that evolution can easily search the entire protein sequence space. Both conclusions are scientifically ridiculous and are inconsistent with what we do understand about proteins…
For the first claim, the evolutionists argue for a smaller protein sequence space because:
A. “the actual identity of most of the amino acids in a protein is irrelevant” and so we can assume there were only a few amino acids in the evolution of proteins, rather than today’s 20.
B. Only the surface residues of a protein are important.
C. Proteins need not be very long. Instead of hundreds of residues, evolution could have used about 50 for most proteins.
For Point A, the evolutionists use as support a series of simplistic studies that replaced the actual protein three-dimensional structure and amino acid chemistries with cartoon, two-dimensional lattice versions.…
Likewise Point B is at odds with science, and again is an unwarranted extrapolation on a simplistic lattice study.
For Point C, the evolutionists note that many proteins are modular and consist of self-contained domains “of as few as approximately 50 amino acids.” But the vast majority of protein domains are far longer than 50 residues. Single domain proteins, and domains in multiple-domain proteins are typically in the hundreds of residues…
At the present time, it is simply an unproven conjecture to suppose that life that was based on only a few amino acids could have originated on Earth, four billion years ago. All living things that we know of require 20 amino acids in order to function. Likewise, it is an unproven conjecture that life-forms requiring only short proteins (less than 50 amino acids long) could ever have existed on Earth. The correct way to justify these conjectures would be to synthesize a life-form that satisfied these conditions. As they say in the state of Missouri, “Show me.” So far, we have seen nothing.
 |
The African hawk-moth Xanthopan morgani. Natural History Museum of London. Image courtesy of Esculapio and Wikipedia.
Why, one might ask, do origin-of-life researchers continue to resort to such arbitrary conjectures? I would suggest that the scientific inspiration for such conjectures comes from an example set by Charles Darwin himself, in 1862, when after viewing an orchid flower with a very long spur to hold its nectar from the island of Madagascar, he made a bold prediction, which was vindicated five years later, that a moth with a proboscis one foot long would be found on that island. Wikipedia takes up the story:
From his observations and experiments with pushing a probe into the spur of the flower, Darwin surmised in his 1862 book Fertilisation of Orchids that there must be a pollinator moth with a proboscis long enough to reach the nectar at the end of the spur. In its attempt to get the nectar at the end of the spur the moth would get pollen rubbed off on its head. The next orchid it visited would then be pollinated in the same manner.
A few years later in 1867 Alfred Russel Wallace published an article in which he supported Darwin’s hypothesis, remarking that the African hawkmoth Xanthopan morganii (then known as Macrosila morganii) had a proboscis almost long enough to reach the bottom of the spur. In a footnote to this article Wallace wrote “That such a moth exists in Madagascar may be safely predicted; and naturalists who visit that island should search for it with as much confidence as astronomers searched for the planet Neptune,–and they will be equally successful!”
It was only in 1903 that a population of Xanthopan morganii (commonly called Morgan’s sphinx moth) with an especially long proboscis was discovered in Madagascar, and it was named subspecies praedicta by Rothschild & Jordan in honor of Wallace’s (not Darwin’s) prediction (Darwin’s prediction was not even mentioned in their paper: Rothschild, L. W. & Jordan, K. 1903. A revision of the Lepidopterous family Sphingidae. Novitates Zoologicae Supplement 9: 1-972). Since Wallace predicted that the mystery pollinator would turn out to be a hawkmoth, rather than simply a large moth as Darwin had suggested in 1903, such a moth was discovered in Madagascar.
It should be noted, however, that Darwin’s and Wallace’s prediction was based entirely on “observations and experiments.” Darwin had every right to predict that a moth with a foot-long proboscis would be found on the island: he had the evidence of a flower with a long spur to back up his prediction.
By contrast, today’s origin-of-life researchers pile supposition upon supposition, without any experimental evidence to back it up. Bold conjectures based on observational evidence are examples of legitimate science, whereas conjectures lacking such evidence are nothing more than castles in the air. Origin-of-life researchers claiming to take their cue from Darwin are therefore making a serious mistake.
A thirteenth fallacy?
Now that I have completed my list of the “dirty dozen,” it occurs to me that there is a thirteenth fallacy: the fallacy of appealing to the progress of science, as grounds for optimism that we’ll eventually solve the problem of how life has originated. Science, we are told, has made great strides in the past, and will continue to do so in the future. An essay titled, Studying the origin of life at the Berkeley University Website Evolution 101 illustrates this kind of thinking:
All the evidence gathered thus far has revealed a great deal about the origin of life, but there is still much to learn. Because of the enormous length of time and the tremendous change that has occurred since then, much of the evidence relevant to origins has been lost and we may never know certain details. Nevertheless, many of the gaps in our knowledge (gaps that seemed unbridgeable just 20 years ago) have been filled in recent years, and continuing research and new technologies hold the promise of more insights. As Ellington puts it, “Origins is a huge knotty problem — but that doesn’t mean it’s an insoluble one.”
The fallacies contained in this argument should by now be readily apparent. First, the argument makes an unwarranted extrapolation: it assumes that scientific progress will continue indefinitely, at the same pace that it has in recent years. Second, the argument makes no attempt to quantify the magnitude of the problem of life’s origin: it may prove to be too large for any conceivable future computer simulation to resolve. Third, the argument naively assumes that the progress of science will serve to diminish the problem, whereas in fact, scientific discoveries of recent years (e.g. the rarity of functional proteins in the space of all possible proteins) have actually served to exacerbate the problem. Thus the problem of life’s origin is far more of a headache for scientists today than it was in Darwin’s day, when the structure of the cell and its chemical constituents remained largely unknown.
To sum up: all the evidence we have to date regarding the origin of life points to its having been intelligently designed. The evidence from proteins is particularly telling in this regard. Only intellectual pigheadedness can account for the reluctance of origin-of-life researchers to discuss the hypothesis that life had an Intelligent Designer, in scientific journals.