The Wikipedia hit-piece on Intelligent Design (NWE’s introductory article is much fairer and better informed) leads with an illustration of a watch; an invidious allusion to William Paley’s famous parable of stumbling over a stone in a field vs. finding a watch in the same field, that appears in Ch I of his 1802 [- 6 ] Natural Theology:
>>IN crossing a heath, suppose I pitched my foot against a stone, and were asked how the stone came to be there, I might possibly answer, that for any thing I knew to the contrary it had lain there for ever; nor would it, perhaps, be very easy to show the absurdity of this answer. But suppose I had found a watch upon the ground, and it should be inquired how the watch happened to be in that place, I should hardly think of the answer which I had before given, that for any thing I knew the watch might have always been there. Yet why should not this answer serve for the watch as well as for the stone; why is it not as admissible in the second case as in the first? . . . >>
He continues:
For this reason, and for no other, namely, that when we come to inspect the watch, we perceive—what we could not discover in the stone—that its several parts are framed and put together for a purpose, e. g. that they are so formed and adjusted as to produce motion, and that motion so regulated as to point out the hour of the day; that, if the different parts had been differently shaped from what they are, or placed after any other manner or in any other order than that in which they are placed, either no motion at all would have been carried on in the machine, or none which would have answered the use that is now served by it.
This is of course, an invitation to the argument by inference to best explanation of empirical observations.
However, it is usually dismissed today as an example of the weakness of attempts to argue by analogy (itself an error, credible analogy is a key facet of real world inductive argument, the cornerstone of science), and the issue put on the table in rebuttal is usually that unlike watches, living things can self-replicate at cellular level, and reproduce with variation, thus evolve. Problem solved, nothing to see here, let us move on.
Not so fast, this dismissal argument is a strawman fallacy.
A strawman?
Yes, Paley’s actual argument has been cut off at a rhetorically convenient point. For, Ch I is titled: “State of the Argument,” and Ch II, “State of the Argument Continued,” and in this chapter (both are fairly short), Paley goes on to discuss precisely the case of a suggested self replicating, time-keeping watch.
Emphasising to draw attention to key steps:
In short, we can see how Paley anticipated Peirce’s reasoning by inference to best explanation by almost a century, and how he raises the key issue of the implications of an object having a functionality and the ADDITIONAL facility of self-replication.
He was of course a generation too early to have the benefit of Babbage’s investigations of digital, stored program computers, but a bar of cams and a stepper to control operations, is just as much a program as anything we have in our computers today. So, he rightly raised the issue that if an object has its own function and the additional facility of replicating itself, that intricately complex additionality demands a proper explanation, one that fits in with what we empirically and analytically know of the credible source of such capabilities.
That brings us to Austrian-American mathematician, John von Neumann, and his 1949 proposal of a kinematic self-replicating machine [vNSR], with the associated issue of the significance of complex, coded, symbolic, algorithmic information in information-processing machines:
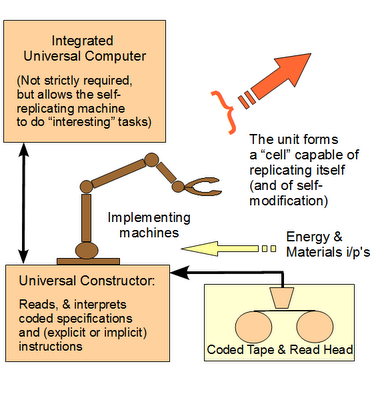
(vi) Also, parts (ii), (iii) and (iv) are each necessary for and together are jointly sufficient to implement a self-replicating machine with an integral von Neumann universal constructor.
(vii) That is, we see here an irreducibly complex set of core components that must all be present in a properly organised fashion for a successful self-replicating machine to exist. [[Take just one core part out, and self-replicating functionality ceases: the self-replicating machine is irreducibly complex (IC).].
(viii) This irreducible complexity is compounded by the requirement (i) for codes, requiring organised symbols and rules to specify both steps to take and formats for storing information, and (v) for appropriate material resources and energy sources.
We can view a ribosome as a degenerate case of [[a Drexler] assembler [[i.e. a molecular scale von Neumann-style replicator]. The ribosome is present in essentially all living systems . . . It is programmable, in the sense that it reads input from a strand of messenger RNA (mRNA) which encodes the protein to be built. Its “positional device” can grasp and hold an amino acid in a fixed position (more accurately, the mRNA in the ribosome selects a specific transfer RNA, which in its turn was bound to a specific amino acid by a specific enzyme). The one operation available in the “well defined set of chemical reactions” is the ability to make a peptide bond [[NB: This works by successively “nudging” the amino acid-armed tip of the codon- matched tRNA in the ribosome’s A site to couple to the amino acid tip of the preceding tRNA (now in the P site) and ratcheting the mRNA forward; thus elongating the protein’s amino acid chain step by step] . . . . [[T]he ribosome functions correctly only in a specific kind of environment. There must be energy provided in the form of ATP; there must be information provided in the form of strands of mRNA; there must be compounds such as amino acids; etc. etc. If the ribosome is removed from this environment it ceases to function.
{kind=link}
{kind=link}
{kind=link}
[[Self Replicating Systems and Molecular Manufacturing, Xerox PARC, 1992. (Parentheses, emphases and links added. Notice as well how the concept of functionally specific complex information naturally emerges from Merkle’s discussion.)]
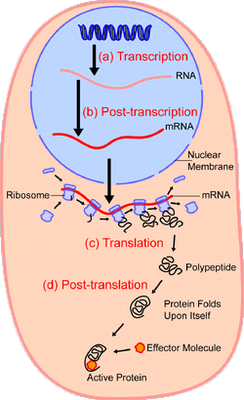
Video (HT BA 77 & UD News):
[vimeo 31830891 Protein Synthesis from Vuk Nikolic]
We are clearly seeing a digital communication system, fitting in the generic framework for such systems:
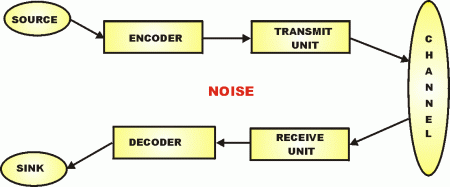
Now, a common attempted rebuttal or dismissal to such reasoning is to claim that spontaneous, natural chance variation [[e.g. through mutations] and natural selection work together to create new functional biological information, so that chance alone does not have to do it all in one step. In Dawkins’ terms, there is alleged to be an easy, step- by- step path up the back slope of “Mt Improbable.” But, this is an error, as quite plainly the claimed source of novel biological information is the variation, not the selection. For, as Darwin himself pointed out in the introduction to Origin, “any being, if it vary however slightly in any manner profitable to itself . . . will have a better chance of surviving, and thus [[will] be naturally selected.”
But the story does not end here.
When we consider the best explanation for complex multicellular organisms organised on the various body plans, we run into the challenge of explaining how a zygote or equivalent replicates itself, then specialises to spatially organise an entity that manifests a complex, tightly functionally integrated body plan; in the case of organisms that undergo metamorphosis, two body plans in a given lifespan.
Meyer, in his 2004 PBSW article (which — contrary to any number of politically correct dismissal attempts — evidently passed proper peer review by “renowned scientists”) highlights why:
In order to explain the origin of the Cambrian animals, one must account not only for new proteins and cell types, but also for the origin of new body plans . . . Mutations in genes that are expressed late in the development of an organism will not affect the body plan. Mutations expressed early in development, however, could conceivably produce significant morphological change (Arthur 1997:21) . . . [but] processes of development are tightly integrated spatially and temporally such that changes early in development will require a host of other coordinated changes in separate but functionally interrelated developmental processes downstream. For this reason, mutations will be much more likely to be deadly if they disrupt a functionally deeply-embedded structure such as a spinal column than if they affect more isolated anatomical features such as fingers (Kauffman 1995:200) . . . McDonald notes that genes that are observed to vary within natural populations do not lead to major adaptive changes, while genes that could cause major changes–the very stuff of macroevolution–apparently do not vary. In other words, mutations of the kind that macroevolution doesn’t need (namely, viable genetic mutations in DNA expressed late in development) do occur, but those that it does need (namely, beneficial body plan mutations expressed early in development) apparently don’t occur.
In short, the concept that we can expect “lucky noise” to write complex, functionally sophisticated, tightly integrated development algorithms for organisms — whether the first cell-based life or more complex multicellular ones including ourselves, is patently a materialist fantasy imposed by redefining science on materialistic premises, not an empirically warranted observation.
And so, Paley’s closing words from Ch I are apt:
Neither, lastly, would our observer be driven out of his conclusion or from his confidence in its truth, by being told that he knew nothing at all about the matter. He knows enough for his argument; he knows the utility of the end; he knows the subserviency and adaptation of the means to the end. These points being known, his ignorance of other points, his doubts concerning other points, affect not the certainty of his reasoning. The consciousness of knowing little need not beget a distrust of that which he does know.
Summing up, it seems that we have a situation where an empirical mechanism for variation and adaptation to niches in environments within existing body plans, has been extrapolated beyond all observational evidence and warranted reason, into a grand mythology of the spo9ntaneous origin of life and of its elaboration based on the alleged powers of lucky noise to write new information incrementally.
The answer to such a priori materialist speculation imposed under the colours of “science” is simple: first, kindly lay aside dismissive, atmosphere-poisoning rhetoric, then empirically demonstrate — with replicability — body plan level macroevolution, on chance variation and differential reproductive success, not philosophical a priorism-loaded extrapolations of variations within the island of function stipulated by an already existing body plan.
Sad to say, after 150 years — despite many confident headlines to the contrary — this challenge has yet to be adequately met. END