By Robert Deyes And John Calvert
PART II: THE ULTIMATE RELATIONSHIP – ANALYZING PATTERNS THAT COMPRISE LIFE
Many scientific disciplines that seek to determine the relationship of an existing pattern to past events analyze them as we analyzed the letters on the drawing board (See PART I). Coroners seek to know the cause of a death – is the death related to a mind or a natural or accidental cause? Those searching for extraterrestrial intelligence seek to know whether a sequence of radio waves from outer space is related to an intelligent rather than a natural or accidental cause. Archeologists seek to know whether a hammer shaped rock got its shape from a mind or a stream.
Our analyses show that the determination of causal relationships involve three inquiries. First, does the pattern manifest a function or purpose – an effect to occur in the future, such as the meaning of the word “Think?” If not there is no necessity to infer a mind. Second, are the various components of the pattern related to or dependent on material causes driven by physical and chemical forces – by necessity? A snowflake looks designed, but its beautiful hexagonal symmetry simply reflects the way water molecules necessarily organize under certain conditions. If chemical necessity can explain the pattern, there is no necessity to infer a mind. Third, if a functional relationship reflected in the pattern is physically independent (not necessary like the snowflake), can chance explain it? If not then a mind – an intention becomes the best explanation for the functional relationship reflected in the pattern. The methodology is explained with great precision by William Dembski in The Design Inference (Refs 1,2).
The methodology for detecting the cause of a pattern is not controversial until one seeks to apply it to the patterns of life. A controversy arises because a conclusion that relates human life to a giver leads to profound implications. There is a difference between a gift and a found object. If one views one’s life as a gift, then it is dependent on the mind of the giver. The recipient might be interested in what the giver wants the gift to be used for. However, if life is just a found object produced by a combination of physical and chemical necessity and chance, then one may do with it as one pleases. If life is a found object it is not dependent on a mind. It is simply the independent result of random interactions of matter, energy and physical forces.
Both conclusions provide the foundation for particular kinds of religion. This is because all religious beliefs depend on an origins narrative that explains the cause of life and its relationship to the world. This was explained by the US Supreme Court in McGowan v. Maryland, 366 U.S. 420 (1961), when it said “religion – in the comprehensive sense in which the Constitution uses that word – is an aspect of human thought and action which profoundly relates the life of man to the world in which he lives.”
Is life a gift or a found object? Is it a creation or the chance occurence (that Jacques Monod comtemplates- see Part I)? Based on belief about the issue, religions then address the subsidiary questions of the nature of life, how life should be lived and what happens when life on earth ends. Traditionals theistic religions explain life as a gift. Non-theistic religions like Atheism and “Secular” Humanism explain it as simply the product of an accumulation of unguided material causes over time. The idea is that life just arises or occurs from materials of the past. These religions hold that life comes from matter, not mind.
Although controversial, investigations of modern science reveal patterns in nature that inexorably lead to the question about how life is related to the world. The data consists of patterns like the word “Think.” Except the patterns that run life are not five letter sequences. They are functional messages that contain billions of symbols not ordered by chemistry. The data is forcing us to think seriously about the adecuacy of chance to provide the answer. Mind is not an absurd alternative for the symphony of function observed in life. Intelligence is ubiquitous to the natural world. We see it in animals, birds, humans and even in cellular systems like the immune system. We are searching for it in outer space. Given the existence of minds, there is warrant for suggesting that our minds may be the product of another mind rather than a chance accumulation of random interactions of matter, energy and physical forces.
The patterns which lead us to a mind are the patterns we observe in DNA. DNA consists of long strands of four different genetic symbols. The genetic symbols are like the symbols Samuel Morse used to carry messages in linear sequences over telegraph lines. Morse used a binary code consisting of dots and dashes. He arbitrarily assigned combinations of dots and dashes to the 26 letters of the alphabet, punctuation marks and numbers. A sequence of three dots “means” the letter “S”. Three dashes “means” the letter “O”.
Like the morse code, the “messages” in DNA are organized per a genetic code of “Eerie Perfection” that uses four kinds of symbols instead of two. Optimization analyses of the genetic code describe it as the best of a million other codes randomly selected from the trillions of possibilities (Ref 3). The four symbols consist of four molecules or nucleotide bases called adenine, cytosine, guanine and thymine. Instead of dots and dashes, the genetic code uses A’s, C’s, T’s and G’s. Like a liner Morse Code message, the messages in DNA are manifested by specific sequences of the four bases along a long linear sugar-phosphate backbone. Two copies of the same message are twined together in the DNA double helix. When it is time for the message to be read (expressed), the two strands are separated and another copy of one stand is made and sent to a translating processor called a ribosome. The ribosome reads the symbols in groups of three symbols called “codons.” Each codon is translated by the ribosome into one of twenty amino acids (like the 26 letters of the English alphabet). The amino acids are then hooked together into long chains that are then folded into three dimensional shapes. The shapes become tools (called enzymes), construction materials (proteins) or operating routines that are used by the cell to build, operate and maintain new cells.
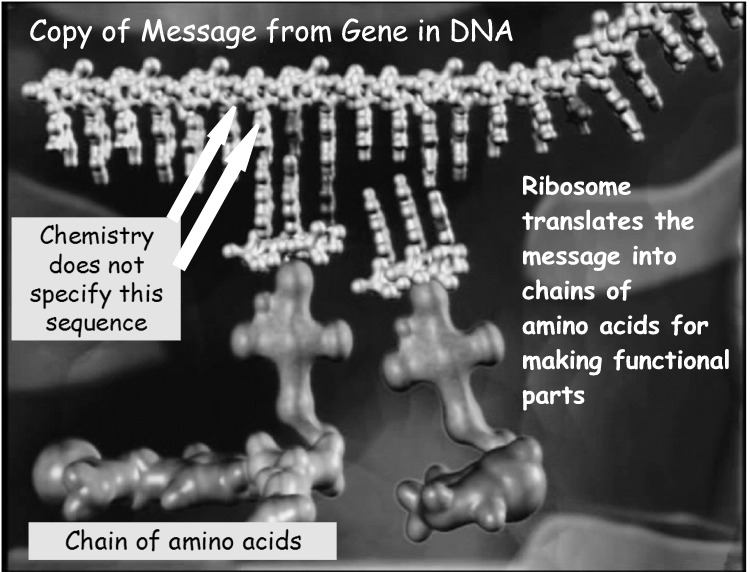
Unlike the square drawing board, DNA is linear, not rectangular. However, the linear strands of DNA are far more extensive than the one hundred squares in the drawing board. It has been estimated that the messages needed to get life started are around 300,000 symbols long (Ref 4). The DNA in each of the 100 trillion cells that comprise a human body is about 3 billion symbols long. If you were to blow up a cell 50,000,000 times its actual size it would be the size of a giant submarine over a mile long. The DNA that is tightly folded in the nucleus would be about 6,000 miles long.
So, how are the genetic symbols in DNA related, if at all? Actually they have the same kinds of relationships as the symbols in the drawing board image that display’s the word “Think.” Each of the “Think” discs are magnetically bound to the board but there is no chemical binding between the discs themselves. Similarly, the nucleotide bases are bound onto the sugar-phosphate backbone of DNA independently of other bases in the DNA sequence. For that reason they may be ordered in any sequence. Stated another way, the genetic symbols, like the letters in the word “Think,” are not related by chemical or physical necessity. If they were, they would not have the capacity to carry the genetic information necessary for life.
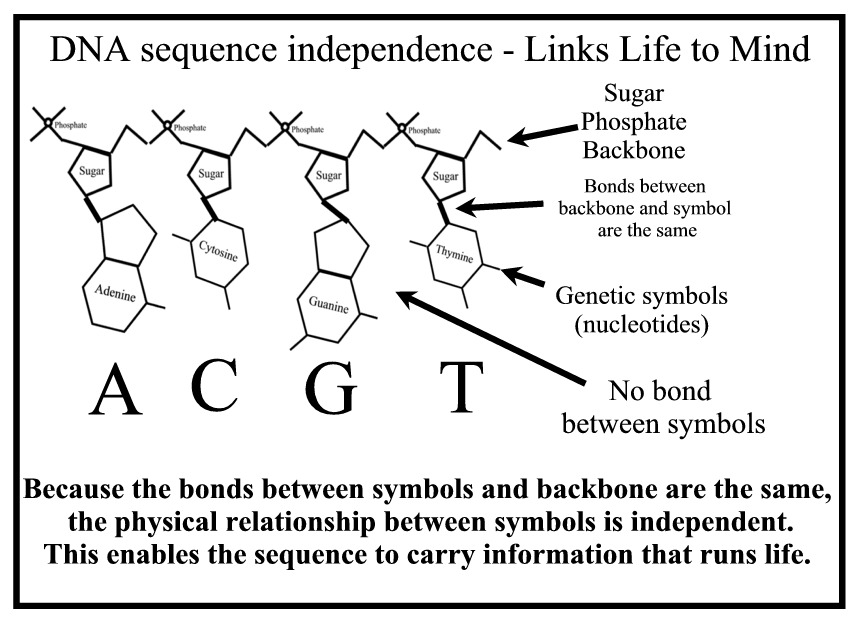
Another relationship that is identical to the “Think” pattern in the vignette we saw in PART I is that the sequence of symbols in DNA is specific for a future function. The meaning of a given DNA message is not hidden like the meaning of the hieroglyphics on the temples at Luxor. Their translation into functional parts, tools and instructions can actually be detected with modern technology. The sequences serve actual, not apparent, future purposes or functions. Like the meaning of the word “Think” the meanings of genetic words, phrases and sentences comprise real information that runs life. It is this functional or semantic characteristic of DNA and all cellular processes that implicates a mind as the cause of life – as the cause of the messages that run life.
Since the nucleotide bases of DNA are not related or ordered by chemistry, the only question that remains is whether chance may plausibly account for those lengthy sequences. Are the messages only “apparent” or illusory messages that are actually attributable to billions of years of random materialistic processes? Even without a statistical analysis, the claim of illusion falls prey to the fact that the messages are actually copied, translated and put to use before our eyes. Statistical analyses simply put the last nail in the coffin of the claim of chance. Like the examples we saw in PART I, the math is compelling because of the exponential decrease in probability.
There are at least 4 possible outcomes (A,C,T or G) at each base position that comprises a genetic message carried by DNA. Getting a specific sequence of 10 bases by chance is not less than 1 over 4x4x4x4x4x4x4x4x4x4 or one over a million (1/410= 1/106). Add another ten bases and the odds drop to about one over a trillion. All plausibility ends with any sequence of over 250 bases.
It takes three bases to specify one of twenty amino acid “letters” in the protein that will eventually be constructed from the translation of the message. These “letter” sequences of amino acids are folded into “words” called proteins or enzymes. The words make up sentences, paragraphs, chapters and whole books of life. An average gene codes for 300 amino acids and therefore consists of about 900 bases. What is the probability of building a gene that codes for a 300 amino acid protein from scratch using our random generator?
T numb. of trials for function(1)
PFprobability of function = ———————————- = 1/10542
O possible outcomes 4900
(Recall that 1 trial = 900 bases; See PART I)
Suppose you have a system that can do one 900 step trial every second. Suppose one starts doing the trials right after the big bang, assuming it occurred a billion times 20 billion years ago, which is about 1025 seconds ago. There are about 32 million seconds in a year. So we can do 1025 trials, if we started work immediately after the big bang. But this still makes the gene unachievable through random means because 1025 divided by 10542 is 1/10517 (this exceeds the Universal Probability Bound 10150 that we discussed earlier).
Suppose we devote more resources to the task. Instead of having only one machine pulling and putting discs, we employ a number of machines equal to every elemental particle in the universe. That is 1080 machines. These resources reduce 1/10517 to 1/10437.
We are now getting closer to probability, but the universal probability bound is not even in sight. What if we had the machines do the trials at the same speed that an elemental particle changes state? That is 1045 per second. We are still far short as 1/10437 minus 1045 = 1/10392.
Now we have a problem. We have reached the limit of probability resources in the entire universe. We are out of time and out of flippers. That is just for one tiny word in a book of information needed for the simplest form of life (Ref 5).
To make matters worse, amino acids degrade very quickly. They are very unstable. So, while we are trying to get the first of 382 genes necessary for life, the environment is constantly switching off the machine. We don’t have billions of years. Maybe we have only an hour. We are like robbers in the bank caught by the police before we have time to run even a few of the trials necessary to get the vault open. The fact that renders the materialistic mechanism impotent is the exponential increase in the amount of probabilistic resources needed for the tasks chance claims to have performed. The exponential increase renders the resources available insufficient. Each additional step in the sequence exponentially increases the time needed to achieve any function, much less all of the function needed to comprise life.
What about Natural Selection – doesn’t that work?
Mind is the most rational explanation for the book-length messages needed to get life started. But suppose we set aside for the moment the question of the origin of life and ask another question. Assume life somehow got started without a mind, can matter alone explain the subsequent library of millions of different books and encyclopedias that contain the messages of the seemingly infinite variety of living forms that presently occupy the Earth?
It is argued that once life arises, a combination of chance and environmental necessity or “natural selection” can sort replicating populations so that only the fit survive. Sometimes populations are split. Each split leads to a new chapter of a new book. Books spawn shelves of books, etc. Eventually the library arises and we are one of the books – a pretty big one.
The inherent problem with the idea is that “natural selection” is not the originator of the new function – new words that comprise the book. The origin of new function remains in the hands of random processes – random changes and additions to an existing book. The environment does not write, it sorts what has been written by chance. It acts as a saboteur until the new function actually arises. New function won’t arise by chance if it is complex because complexity requires more time than is available to a random process.
For example, suppose the first form of life is a bacteria that cannot swim towards a food source. It lives only if it is served by an environmental waitress. It would be more fit for a cafeteria environment if it could swim to the food bar. To achieve that new “function” it needs a new series of genetic instructions that will assemble, operate and maintain a motor, systems to deliver energy to the motor, systems designed to sense food in the environment and systems that will steer the moving organism to the food. Suppose one component of the motor consists of a new kind of bushing. The motor won’t work without the bushing. A new gene is needed to specify the recipe for building a component of the bushing, another is needed to specify the time it is to be built, another is needed to specify how many are to be built, another is needed to take it to the right place in three dimensional space where it is to be used, and another is needed to cause it to be incorporated into the structure that may or may not ever arise.
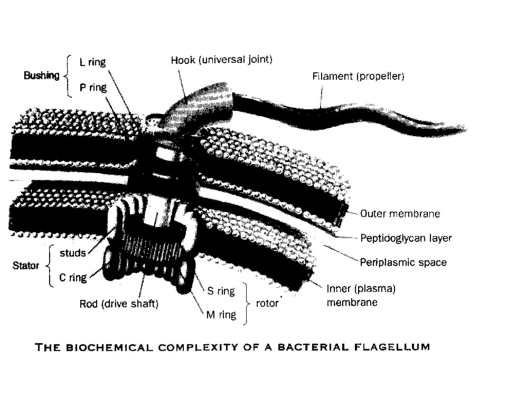
Suppose the new gene for the recipe of the bushing requires a sequence of 900 nucleotide bases before its “function” as a part for a bushing arises. Until that function arises, natural selection does not act to save it. It more probably deletes the energy robbing proto gene before even a short part of it is ever formed. In fact, there is not enough time for a random process to produce a gene that requires a specific sequence of 900 nucleotides. But even if the miracle happens, the organism still can’t swim to the food bar. Once the new gene arises, other components in the cell must also be present to transcribe and ferry the bushing to where it is needed at the right time. But, the motor requires more than just a bushing. It requires many more components.
In the end, chance is just as much of a problem for increases in the diversity of life as it is for the origin of life itself. Natural selection may do a reasonable job of fine-tuning an existing population, but it is not the friend of innovation.
The primary defect of the unobserved hypothesized process of biological evolution is the absurd implausibility of the claim that a random mechanism can produce the sophisticated array of functional systems needed to run life. The exponential increase in the time necessary for each new step needed to attain the required function is the killer. Like a house of cards, the assembly of machines themselves requires an orchestrated timing. One cannot start building a sand castle today and expect to finish the job a year later after natural selection has torn it down.
A paper by biochemist Michael Behe and mathematician David Snoke illustrates the problem with the process even where new function is derived by randomly changing genetic letters in an existing duplicate gene rather than the development of an entirely new gene (Ref 6). It is thought that much biological diversity arises from accidental duplication of genes, where the duplicate serves no apparent function in the genome. The speculation is that as the duplicate gene is randomly mutated new positive functions may arise that will make the organism more fit. However, new function often requires multiple changes, not just one. It may take as many as thirteen steps to get the new function. The Behe – Snoke paper shows that to get new function requiring only six changes, in a replicating population of bacteria one would need to have a population the size of 1021 (one trillion, trillion, trillion, trillion) replicating over 100 million generations.
New discoveries about DNA have caused scientists to question the adequacy of chance to explain biological function. A 2006 paper concludes that 80,000 “simple sequence repeats,” or “SSR’s” found in the human genome are not likely due to chance. SSR’s are segments of DNA not contained in genes that previously were thought to be non functional “junk.” They are short sequences of bases that on average repeat more than 50 times in a series, such as “CTGCAG CTGCAG CTGCAG…..” The author explains: “The probability that a particular sequence of n base pairs will appear at a specified site in a random DNA sequence is approximately (1/4)n [assuming equal proportions of each nucleotide]. Thus any repeated sequence longer than 20 or so base pairs is unlikely to appear solely by chance, even once, anywhere in the 3×109 base pairs of the human genome.)” (Ref 7). If an SSR has an average length of 6 bases and each sequence has an average length of 50 repeats, then the average length of just one of the 80,000 SSR’s is 300 bases. Thus the probability of a chance formation of one SSR is 1/4300 or roughly 1/10180 or practically zero.
The problem of chance also arises in the context of gene expression. New discoveries have shown that genes may be turned on or off and thereby made to combine selectively with a repertoire of other genes that have been turned on or off. A dictionary contains thousands of words. Similarly, the genome contains thousands of genes or genetic “words.” Just as every word in the dictionary can be used or not used in a variety of combinations to express thoughts, the same is apparently true of genes in the genome. Stuart Kaufman, a scientist seeking to develop a new theory of evolution, believes that differences in morphology may be due to the sequence in which genes are used or not used. He notes that the possible combinations are enormous: “Well there’s 25,000 genes, so each could be on or off. So there’s 2x2x2x25,000 times. Well that’s 2 to the 25,000th. Right? Which is something like 10 to the 7,000th. Okay? There’s only 10 to the 80th particles in the whole universe. Are you stunned?” (Ref 8 ).
Accordingly, random events and random mutations are not good explanations for the messages needed for life. Since the messages are not related by chance or necessity, their clearly evident function and purpose inexorably leads one to the mind of an author.
Just as one cannot believe that the word “Think” appeared without the help of the sneaky mind of the intellectually gifted Maggie (See Part I), one is forced to look to a mind to explain life. Indeed the activity of that mind is not really sneaky at all. Its products are easy for everyone to see. We really have no rational excuse for denying it.
ABOUT THE AUTHORS
Robert Deyes holds a Masters Degree in Medical Genetics from the University of Glasgow and a Bachelors Degree in Molecular Biology from the University of Portsmouth, UK. He carried out an 18-month research project at the Université Louis Pasteur in Strasbourg, France.
John Calvert, JD, holds a Bachelors degree and professional experience in Geology and is Managing Director of Intelligent Design Network Inc, a non-profit organization that seeks institutional objectivity in origins science.
BIBLIOGRAPHY
1. William Dembski in The Design Inference: Eliminating Chance Through Small Probabilities, p.36 et. seq (Cambridge Press, 1998)
2. William Dembski in No Free Lunch: Why Specified Complexity Cannot Be Purchased Without Intelligence, p.83 (Rowman & Littlefield, 2002)
3. S.J. Freeland and L.D. Hurst, The Genetic Code is One in Million, (Journal of Molecular Evolution, vol. 47, pp 238-248, 1998); In Life’s Solution: Inevitable Humans in a Lonely Universe, p.13-21, (Cambridge Press, 2003) paleontologist Simon Conway Morris devotes a sub-chapter to the extraordinary efficiency of the Genetic Code. He captions it “EERIE PERFECTION.”
4. John Glass, et. al., Essential genes of a minimal bacterium, Synthetic Biology Group, J. Craig Venter Institute, (Proceedings of the National Academy of Science, Jan 2006, vol 103, No. 2, 425-430). The study concluded that the simplest form of life known to have existed requires around 382 “essential” genes. The average length of a gene of a prokayote is around 900 nucleotides, or 343,800 nucleotides for the 382 essential genes.
5. Comparing the probability of the Amino Acid Sequence with the probability of the DNA sequence: Each codon or sequence of three nucleotides in DNA is translated into one amino acid. The acids are hooked together into a long chain that is then folded into a three dimensional shape. The shape of the protein determines its function. There are only 20 amino acids that are used and therefore the probability of any amino acid being in a particular position in the string arguably is 1/20. If the string is 300 amino acids long, the probability is 1/20300 or 1/10390. This is a higher probability then the probability of the DNA sequence that specifies the amino acids in the string. That probability is ¼ 900 or 1/ 10542. The reason for the difference is that a number of the 64 possible nucleotide codons specify for the same amino acid. This reduces the effect of errors in copying and translation. However, the difference in probability is actually insignificant because any complex sequence will exhaust all available probabilistic resources very quickly.
6. Michael Behe & David Snoke, Simulating evolution by gene duplication of protein features that require multiple amino acid residues (Protein Science, The Protein Society September 2, 2004)
7. David King, Edward Trifonov and Yechezkel Kashi, Tuning Knobs in the Genome: Evolution of Simple Sequence Repeats by Indirect Selection published in The Implicit Genome, Lynn Caporale, Editor, p 77 (Oxford University Press, 2006)
8. Suzan Mazur, The Altenberg 16: Will the Real Theory of Evolution Please Stand Up? (Scoop Independent News, July 2008), the online edition is posted at http://www.scoop.co.nz/stories/HL0807/S00053.htm. A conference was held in Altenberg, Austria in July 2008, for the purpose of developing a new theory of evolution because the current one does not fit the data.