This lecture by Dr Stephen Meyer of Discovery Institute, with Q & A may be a good refresher and focus for thought on OoL, HT WK:
WK — a useful blog to bookmark and monitor to see trends and issues — gives a helpful bullet point outline, in part:
- intelligent design is concerned with measuring the information-creating capabilities of natural forces like mutation and selection
- Darwinists think that random mutations and natural selection can explain the origin and diversification of living systems
- Darwinian mechanisms are capable of explaining small-scale adaptive changes within types of organisms
- but there is skepticism, even among naturalists, that Darwinian mechanisms can explain the origin of animal designs
- even if you concede that Darwinism can account for all of the basic animal body plans, there is still the problem of life’s origin
- can Darwinian mechanisms explain the origin of the first life? Is there a good naturalistic hypothesis to explain it?
- there are at least two places in the history of life where new information is needed: origin of life, and Cambrian explosion . . .
- it is very hard to arrive at a meaningful sequence of a non-trivial length by randomly picking symbols/letters
- although any random sequence of letters is improbable, the vast majority of sequences are gibberish/non-compiling code
- similarly, most random sequences of amino acids are lab-proven (Doug Axe’s work) to be non-functional gibberish
- the research showing this was conducted at Cambridge University and published in the Journal of Molecular Biology
- so, random mutation cannot explain the origin of the first living cell
- however, even natural selection coupled with random mutation cannot explain the first living cell
- there must already be replication in order for mutation and selection to work, so they can’t explain the first replicator
- but the origin of life is the origin of the first replicator – there is no replication prior to the first replicator
- the information in the first replicator cannot be explained by law, such as by chemical bonding affinities . . . .
- if laws did determine the sequence of letters, then the sequences would be repetitive
- the three materialist explanations – chance alone, chance and law, law alone – are not adequate to explain the effect
- the best explanation is that an intelligent cause is responsible for the biological explanation in the first replicator
Where, of course, there is only one empirically observed, directly known causal factor capable of causing functionally specific complex organisation and associated information, especially digitally coded functionally specific complex information, design.
Clipping some key points:
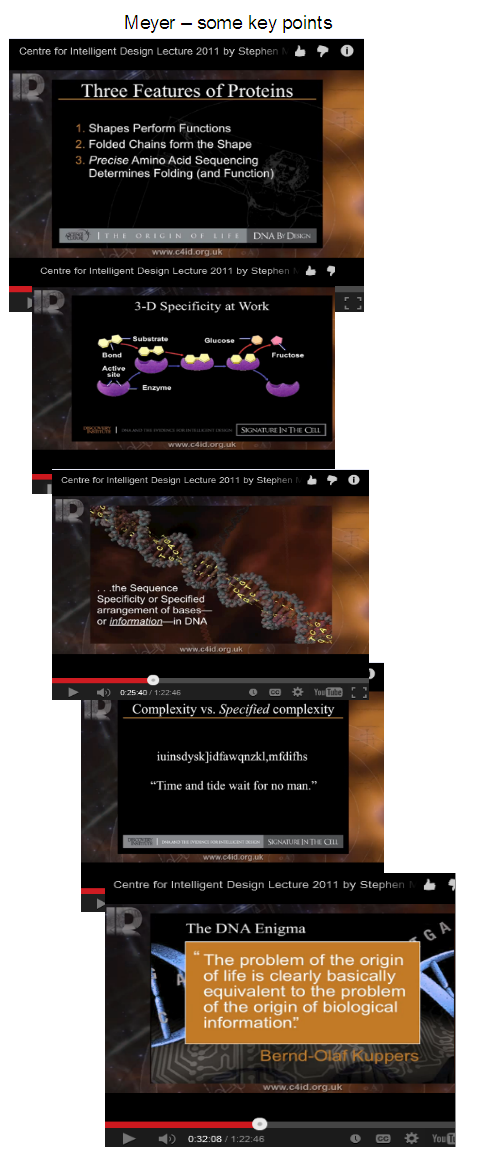
{U/D, Jan 10:} It is also useful to follow a chain of Meyer’s argument to the inference to design:
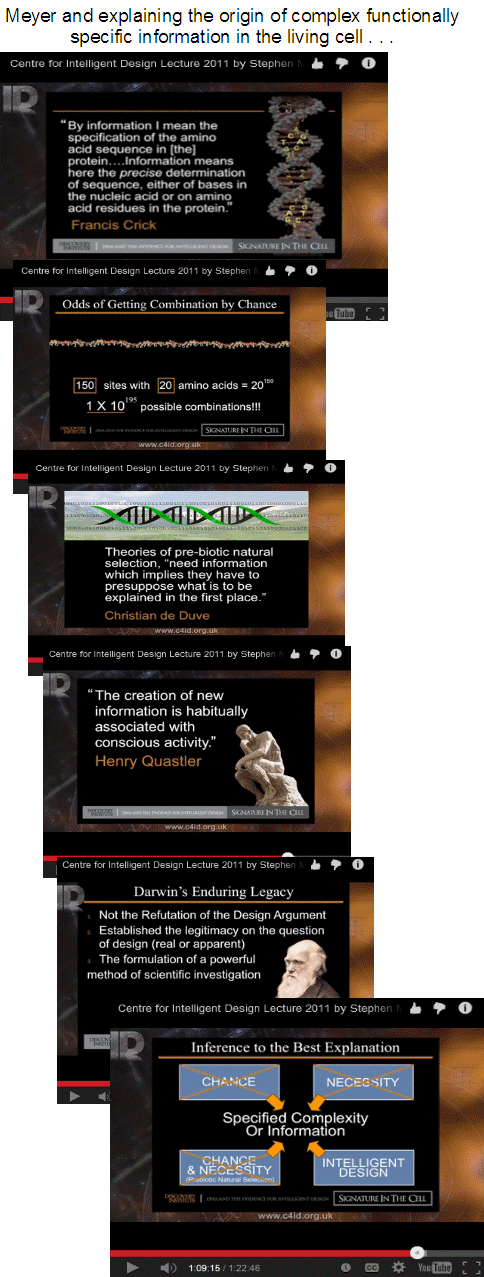
So we see here an inductive inference to best explanation as a method of historical/origins sciences, leading to the conclusion that phenomena or objects manifesting functionally specific, complex organisation and associated information (FSCO/I) are best explained, not on blind chance and/or mechanical necessity, but on intelligent design.
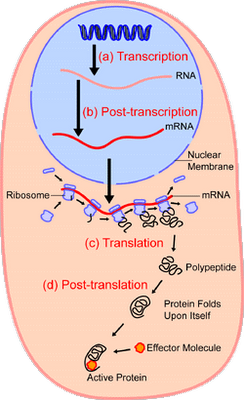
Recall, too, the cell’s basic functionality includes the protein synthesis chain, which is patently a case where all core steps and components are required in proper order — irreducible complexity, in short — for the system to function:
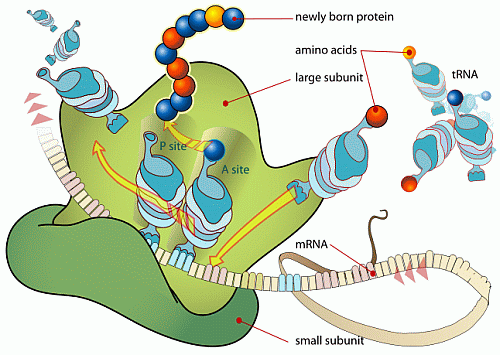
. . . which also includes the digital information controlled, algorithmic assembly process in the Ribosome:
. . . as well as fulfilling Mignea’s irreducibly complex requisites for self replicating life:
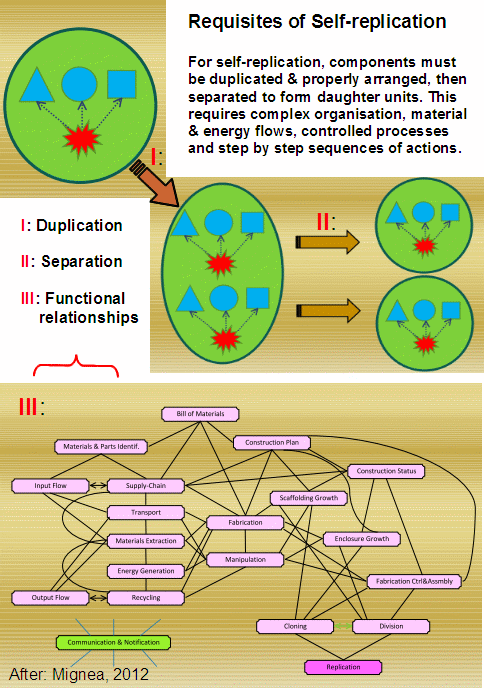
( See Mignea, 2012, slide show; fair use. Presentation speech is here.)
. . . with of course the force of the design inference on high contingency joined to complexity and to specificity to function — especially functionally specific complex organisation and associated information beyond 500 – 1,000 bits [FSCOI] also involved in our thinking:
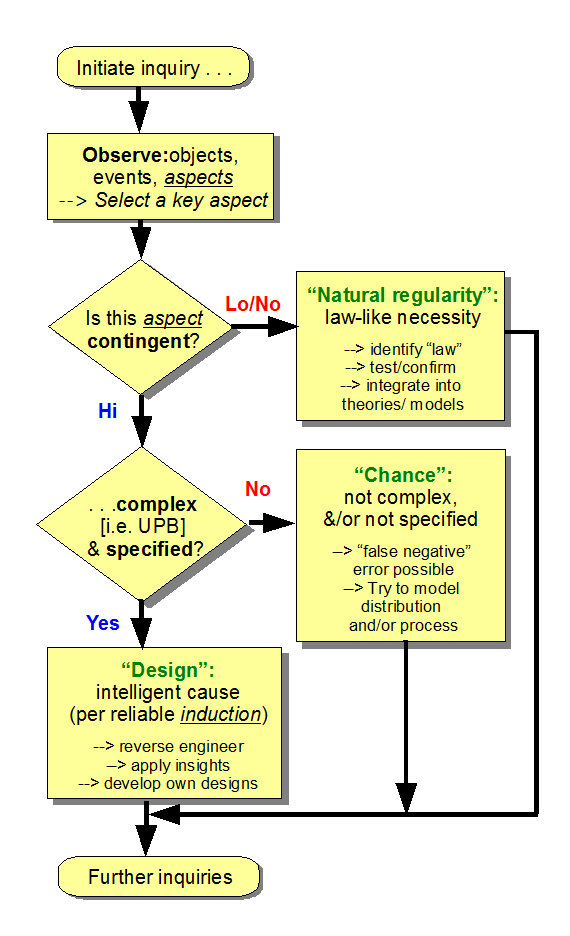
(As in can you kindly provide a clear case of FSCO/I arising by blind chance and mechanical necessity in our actual observation? And just what is the routinely seen cause of such FSCO/I? So, then on the requirement of candidate explanations being actually observed causes, and like causes like, what best explains OOL? Why?)
So, where does inference to best, empirically grounded inductive explanation point? Why? END
PS: I strongly urge actually watching the Meyer video as a basis for discussion.