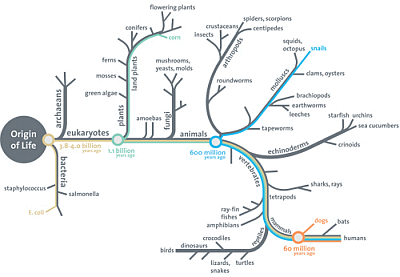
In recent exchanges in and around UD on origins and the tree of life, Theobald’s 29 evidences claims (and by implication the sort of summary presented by Wikipedia in its articles on Abiogenesis and Evolution) have come up. [NB: to carry forward discussions, I suggest here on. I intend to do a for reference in support of discussion here in this FTR post.]
That leads me to point out the case of the UD pro-darwinism essay challenge and the strange absence of and reluctance to provide a guest essay here at UD over the course of a full year, Sept/Oct 2012 – Sept/Oct 2013. The big issue seemed to be that in my challenge as explained, I required tackling the whole tree of life from its root in OOL on up (on grounds that this is the heart of the common origin of FSCO/I challenge).
Eventually, with Darwinist objectors stung by their lack of ability to take a free kick at goal, I was able to piece together a composite — but sadly inadequate — reply.
It is revealing that — after a full year to address the matter — the OOL side of the issue had to be addressed as follows:
PART I:
____________
EL, 149: >> “As yet we have no empirically supported naturalistic theory of abiogenesis.” >>
____________
It hardly needs to be pointed out regarding this case, that FSCO/I — a main feature of cell-based life forms — has one well known, routinely observed source: design.
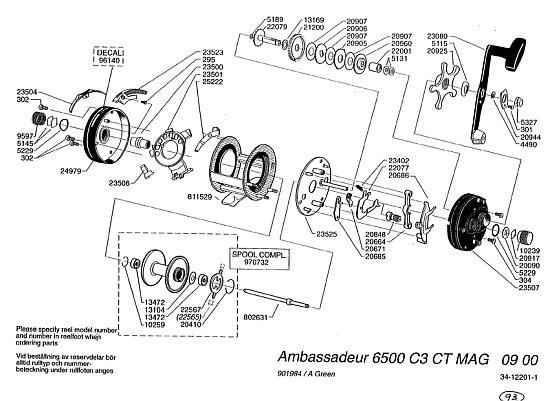
A point that is closely tied to how the requisites of specific functionality based on interaction of many, correctly arranged components forces the right parts to be arranged in the right nodes and arcs pattern to achieve function, e.g. in the classic ABU 6500 3c reel:
. . . or, in the process-flow wiring diagram patterns of a petroleum refinery:

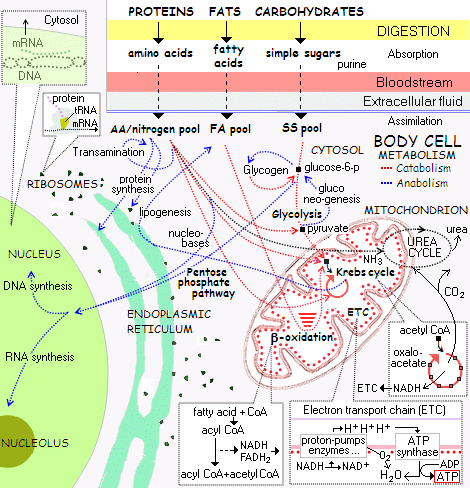
. . . which closely parallel those of cellular metabolism (for much more detail cf here):
. . . which lead to the classic challenge of finding islands of function in large configuration spaces through blind search based on chance and mechanical necessity:
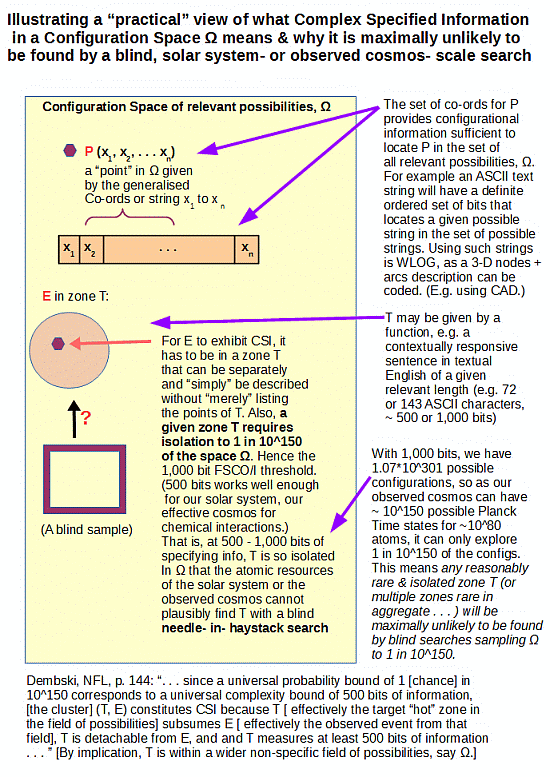
. . . in a further context of the lack of resources to effect thorough search imposed by the atomic resources and time accessible to the solar system (or the observed cosmos):
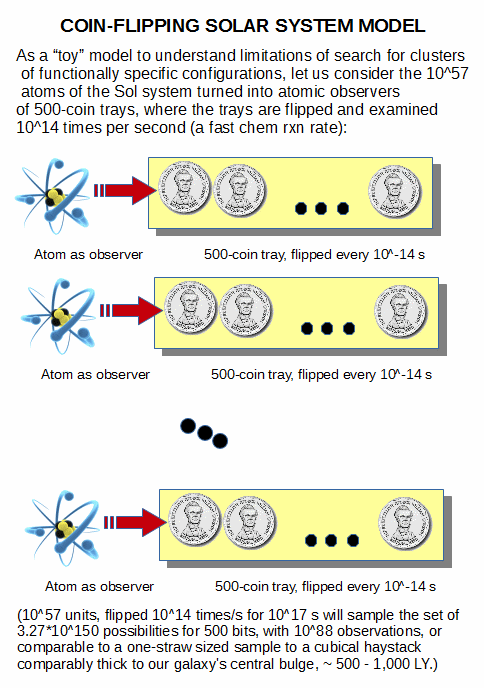
. . . where the real FSCO/I generation problem to get to cell based life starts with get a plausible Darwin’s warm pond of salts or the like and with reference only to blind physics and chemistry show how in a way that can be backed up by empirical observation, we credibly end up with cell based life with genetic codes, metabolisms, proteins as workhorse molecules, protein assembly machines and smart gating of protective enclosures.
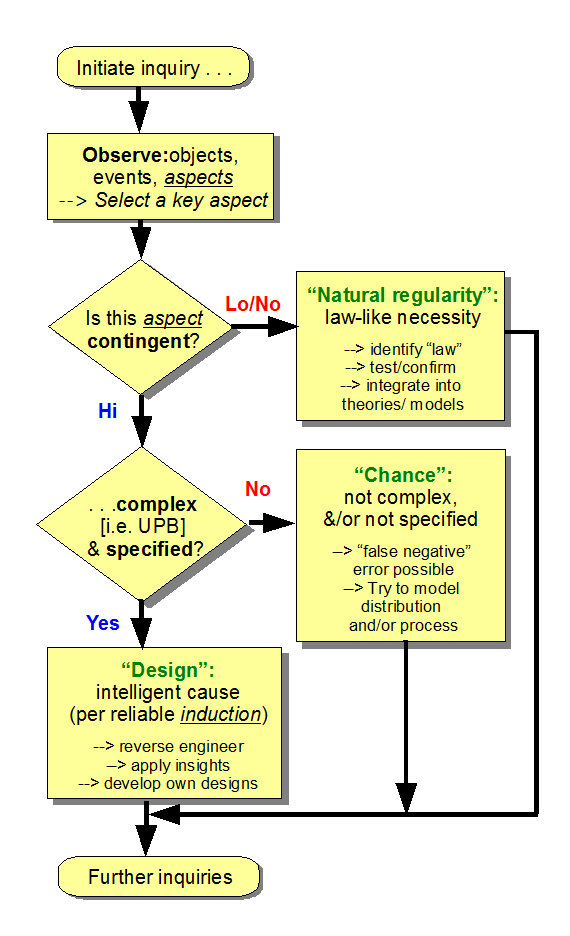
{U/D Nov 7:} Where, we may summarise the design inference process through a flowchart that shows how if blind chance and/or mechanical necessity can and in our observation do account for the sort of features and aspects in view, those are accepted as adequate explanation. Only on seeing that combination of specified complexity that is often tied to multi-part function that depends on proper organisation and interaction of many parts do we infer design based on the relevant inductive base that shows that for instance FSCO/I is routinely, reliably and only known per observations to come about by intelligently directed configuration, aka design:
For more specific biological example (HT: Wiki) we may observe the protein assembly process (knowing that proteins are the workhorse molecules of the living cell:
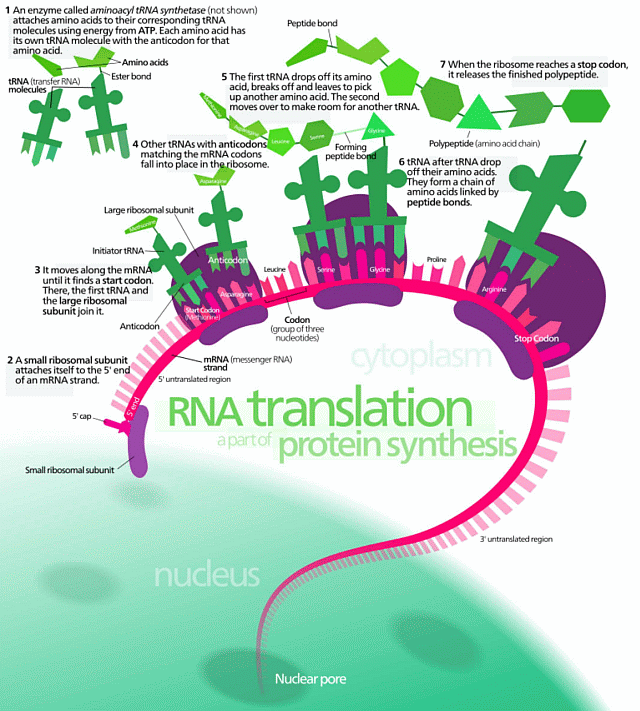
vid:
[vimeo 31830891]
And make some useful comparisons on the FSCO/I involved:

On fair comment, the other half of the composite response was little better, on the challenge of accounting per naturalistic models, for the FSCO/I involved in origin of major body plans. My key responsive remark therefore began:
First, we see:
[KF:] As the main point in the OP highlights, there is but one
empirically justified source of FSCO/I, design. That is, empirically
and inductively, FSCO/I is known to be a reliable sign of design, as
deer tracks are reliable signs of the passage of certain animals.
[J:] As usual, I beg to differ. At least as far as the
diversification of life on earth is concerned given a as yet
unknown original replicator.
The fact remains, that the only actually observed source
of FSCO/I is indeed design. You have substituted an inadequately
grounded assertion as though it were a factual observation to the
contrary. Unfortunately, as Theobald indicates, this confusion of a
tower of inferences for indisputable facts we have observed is a
characteristic problem of evolutionary materialist thought.
You have not seen not even a remainder of that hypothetical
replicator and lush documentaries etc to the contrary, we do not have
observations of the incrementalist origin of body plans either . . .
[u/d:Prof Philip Johnson lectures on the logical issues with Darwinism:]
[youtube gwj9h9Zx6Mw]
But also, during the process of making and addressing the challenge, Wikipedia and Theobald’s 29+ evidences were put to sit in the otherwise empty chair . . . empty until the very unsatisfactory composite was put into play.
First, Wiki:
_________________
Root: >>Article on Abiogenesis:
>>Abiogenesis . . . or biopoiesis is the study of how biological life could arise from inorganic matter through natural processes. In particular, the term usually refers to the processes by which life on Earth may have arisen. Abiogenesis likely occurred between 3.9 and 3.5 billion years ago, in the Eoarchean era (i.e. the time after the Hadean era in which the Earth was essentially molten).
Hypotheses about the origins of life may be divided into several categories. Most approaches investigate how self-replicating molecules or their components came into existence. For example, the Miller–Urey experiment and similar experiments demonstrated that most amino acids, often called “the building blocks of life”, were shown to be racemically [–> i.e. no answer for handedness] synthesized in conditions thought to be similar to those of the early Earth. [–> Of course this is highly disputable] Several mechanisms have been investigated, including lightning and radiation. Other approaches (“metabolism first” hypotheses) focus on understanding how catalysis in chemical systems in the early Earth might have provided the precursor molecules necessary for self-replication . . . .
There is no “standard model” of the origin of life. Most currently accepted models draw at least some elements from the framework laid out by the Oparin-Haldane hypothesis. Under that umbrella, however, are a wide array of disparate discoveries and conjectures . . . >>
So, OOL is a major challenge to a would-be essayist.>>
Branch: >> Let’s continue to survey and critique the modern grand blind watchmaker evolutionary synthesis, in outline. (Looking at the overall summary allows us to see the logical patterns and issues as we look at the forest as a whole and don’t get lost in the trees. I suspect this is where a lot of the reluctance to provide a 6,000 word survey comes from. Never mind, we can pull up a handy summary of conventional wisdom substitute for the empty chair we are facing.)
Back to Wiki standing in, here on evolution (but we must first recall from yesterday as already clipped, that there is no sound answer to the OOL challenge so the Darwinist tree of life has no root):
Evolution is the change in the inherited characteristics of biological populations over successive generations. Evolutionary processes give rise to diversity at every level of biological organisation, including species, individual organisms and molecules such as DNA and proteins.[1]
Life on Earth originated and then evolved from a universal common ancestor approximately 3.7 billion years ago. Repeated speciation and the divergence of life can be inferred from shared sets of biochemical and morphological traits, or by shared DNA sequences. These homologous traits and sequences are more similar among species that share a more recent common ancestor, and can be used to reconstruct evolutionary histories, using both existing species and the fossil record. Existing patterns of biodiversity have been shaped both by speciation and by extinction.
Charles Darwin was the first to formulate a scientific argument for the theory of evolution by means of natural selection. [–> Wallace the ID-ish heretic gets left out . . . ] Evolution by natural selection is a process that is inferred from three facts about populations: 1) more offspring are produced than can possibly survive, 2) traits vary among individuals, leading to differential rates of survival and reproduction, and 3) trait differences are heritable.[3] Thus, when members of a population die they are replaced by the progeny of parents that were better adapted to survive and reproduce in the environment in which natural selection took place. This process creates and preserves traits that are seemingly fitted for the functional roles they perform.[4] Natural selection is the only known cause of adaptation, but not the only known cause of evolution. Other, nonadaptive causes of evolution include mutation and genetic drift.[5]
In the early 20th century, genetics was integrated with Darwin’s theory of evolution by natural selection through the discipline of population genetics. The importance of natural selection as a cause of evolution was accepted into other branches of biology. Moreover, previously held notions about evolution, such as orthogenesis and “progress” became obsolete.[6] Scientists continue to study various aspects of evolution by forming and testing hypotheses, constructing scientific theories, using observational data, and performing experiments in both the field and the laboratory. Biologists agree that descent with modification is one of the most reliably established facts in science.[7] Discoveries in evolutionary biology have made a significant impact not just within the traditional branches of biology, but also in other academic disciplines (e.g., anthropology and psychology) and on society at large . . . .
In the 1920s and 1930s a modern evolutionary synthesis connected natural selection, mutation theory, and Mendelian inheritance into a unified theory that applied generally to any branch of biology. The modern synthesis was able to explain patterns observed across species in populations, through fossil transitions in palaeontology, and even complex cellular mechanisms in developmental biology.[23][42] The publication of the structure of DNA by James Watson and Francis Crick in 1953 demonstrated a physical basis for inheritance.[43] Molecular biology improved our understanding of the relationship between genotype and phenotype. Advancements were also made in phylogenetic systematics, mapping the transition of traits into a comparative and testable framework through the publication and use of evolutionary trees.[44][45] In 1973, evolutionary biologist Theodosius Dobzhansky penned that “nothing in biology makes sense except in the light of evolution”, because it has brought to light the relations of what first seemed disjointed facts in natural history into a coherent explanatory body of knowledge that describes and predicts many observable facts about life on this planet.[46]
Since then, the modern synthesis has been further extended to explain biological phenomena across the full and integrative scale of the biological hierarchy, from genes to species. This extension has been dubbed “eco-evo-devo” . . . .
Evolution by means of natural selection is the process by which genetic mutations that enhance reproduction become and remain, more common in successive generations of a population. It has often been called a “self-evident” mechanism because it necessarily follows from three simple facts:
Heritable variation exists within populations of organisms.
Organisms produce more progeny than can survive.
These offspring vary in their ability to survive and reproduce.These conditions produce competition between organisms for survival and reproduction. Consequently, organisms with traits that give them an advantage over their competitors pass these advantageous traits on, while traits that do not confer an advantage are not passed on to the next generation.[93]
The central concept of natural selection is the evolutionary fitness of an organism.[94] Fitness is measured by an organism’s ability to survive and reproduce, which determines the size of its genetic contribution to the next generation.[94] However, fitness is not the same as the total number of offspring: instead fitness is indicated by the proportion of subsequent generations that carry an organism’s genes.[95] For example, if an organism could survive well and reproduce rapidly, but its offspring were all too small and weak to survive, this organism would make little genetic contribution to future generations and would thus have low fitness.[94] . . . .
Evolution influences every aspect of the form and behaviour of organisms. Most prominent are the specific behavioural and physical adaptations that are the outcome of natural selection. These adaptations increase fitness by aiding activities such as finding food, avoiding predators or attracting mates. Organisms can also respond to selection by co-operating with each other, usually by aiding their relatives or engaging in mutually beneficial symbiosis. In the longer term, evolution produces new species through splitting ancestral populations of organisms into new groups that cannot or will not interbreed.
These outcomes of evolution are sometimes divided into macroevolution, which is evolution that occurs at or above the level of species, such as extinction and speciation and microevolution, which is smaller evolutionary changes, such as adaptations, within a species or population.[141] In general, macroevolution is regarded as the outcome of long periods of microevolution.[142] Thus, the distinction between micro- and macroevolution is not a fundamental one – the difference is simply the time involved.[143] . . . .
Highly energetic [= thermodynamically unfavourable, hence hard to account for spontaneously in plausible prebiotic environments] chemistry is thought to have produced a self-replicating molecule around 4 billion years ago and half a billion years later the last common ancestor of all life existed.[241] The current scientific consensus [–> appeal to authority and to no true scotsman, where in fact a major debate and conundrum is being papered over] is that the complex biochemistry that makes up life came from simpler chemical reactions.[242] The beginning of life may have included self-replicating molecules such as RNA,[243] and the assembly of simple cells.[244] [–> restating the problem with some speculations inserted, as though it is the solution] . . . .
All organisms on Earth are descended from a common ancestor or ancestral gene pool. [–> begging the question] [173][245] Current species are a stage in the process of evolution, with their diversity the product of a long series of speciation and extinction events.[246] The common descent of organisms was first deduced from four simple facts about organisms: First, they have geographic distributions that cannot be explained by local adaptation. Second, the diversity of life is not a set of completely unique organisms, but organisms that share morphological similarities. Third, vestigial traits with no clear purpose resemble functional ancestral traits and finally, that organisms can be classified using these similarities into a hierarchy of nested groups – similar to a family tree.[247] However, modern research has suggested that, due to horizontal gene transfer, this “tree of life” may be more complicated than a simple branching tree since some genes have spread independently between distantly related species. [–> burying the evidence of mutually incompatible molecular trees and the evidence of code libraries under an assertion; U/D Nov 5, 2014: I now add, and so much for nested hierarchies which necessarily show up as branching trees . . . U/B Nov 6, cf here for an in-thread comment giving a few details in reply to KS ] [248][249]
Past species have also left records of their evolutionary history. Fossils, along with the comparative anatomy of present-day organisms, constitute the morphological, or anatomical, record.[250] By comparing the anatomies of both modern and extinct species, paleontologists can infer the lineages of those species. [–> U/D Nov 5, 2014, on what assumptions? with what empirical support per observations on cause of FSCO/I? How is circular, question begging avoided, especially given claims of self evidence highlighted above, which are patently unwarranted.] However, this approach is most successful for organisms that had hard body parts, such as shells, bones or teeth. Further, as prokaryotes such as bacteria and archaea share a limited set of common morphologies, their fossils do not provide information on their ancestry.
More recently, evidence for common descent has come from the study of biochemical similarities between organisms. For example, all living cells use the same basic set of nucleotides and amino acids.[251] The development of molecular genetics has revealed the record of evolution left in organisms’ genomes: dating when species diverged through the molecular clock produced by mutations.[252] For example, these DNA sequence comparisons have revealed that humans and chimpanzees share 96% of their genomes and analyzing the few areas where they differ helps shed light on when the common ancestor of these species existed.[253] [–> Where of course this very case is riddled with all sorts of highly questionable assumptions and assertions] . . . .
The history of life was that of the unicellular eukaryotes, prokaryotes and archaea until about 610 million years ago when multicellular organisms began to appear in the oceans in the Ediacaran period.[255][262] The evolution of multicellularity occurred in multiple independent events, in organisms as diverse as sponges, brown algae, cyanobacteria, slime moulds and myxobacteria.[263]
Soon after the emergence of these first multicellular organisms, a remarkable amount of biological diversity appeared over approximately 10 million years, in an event called the Cambrian explosion. Here, the majority of types of modern animals appeared in the fossil record, as well as unique lineages that subsequently became extinct. [[–> top down, not bottom up . . . what is this telling us?] [264] Various triggers for the Cambrian explosion have been proposed, including the accumulation of oxygen in the atmosphere from photosynthesis.[265]
About 500 million years ago, plants and fungi colonised the land and were soon followed by arthropods and other animals.[266] Insects were particularly successful and even today make up the majority of animal species.[267] Amphibians first appeared around 364 million years ago, followed by early amniotes and birds around 155 million years ago (both from “reptile”-like lineages), mammals around 129 million years ago, homininae around 10 million years ago and modern humans around 250,000 years ago.[268][269][270] However, despite the evolution of these large animals, smaller organisms similar to the types that evolved early in this process continue to be highly successful and dominate the Earth, with the majority of both biomass and species being prokaryotes.[151] . . .
There is of course no discussion of the origin of functionally specific biological information challenge, there is an obvious assumption that incremental variation is sufficient to explain all of biodiversity, and the OOL challenge is severely understated.
Notice in particular the notion that macro-evo is simply cumulative micro-evo.
That is a strong reflection of the assumption of incrementalism.
One of the most stunning bland false assertions above is this, and it is a doozy:
Natural selection is the only known cause of adaptation
We live in a world with literally billions of observed incidents of adaptation by design. This is an unwarranted and false assertion that plays a material part in deciding the issue before the facts can speak.
The definition and discussion of the power of natural selection cleverly skirts the problem of a claimed but dubious self-evident truth: grand question-begging.
A true self evident truth is one that is seen as such once one understands what is asserted, on pain of patent absurdity on attempted denial, e.g. error exists. What is really happening here is that this is a major assumption not open to question in the system and seen as being so obviously “true” that if you question you are perceived as being an outsider and fair game for dismissal. After all if something is self evident then only one who insists on absurdities will reject it.
But in fact a closer look will reveal a major gap:
Evolution by means of natural selection is the process by which genetic mutations that enhance reproduction become and remain, more common in successive generations of a population. It has often been called a “self-evident” mechanism because it necessarily follows from three simple facts:
Heritable variation exists within populations of organisms.
Organisms produce more progeny than can survive.
These offspring vary in their ability to survive and reproduce.These conditions produce competition between organisms for survival and reproduction. Consequently, organisms with traits that give them an advantage over their competitors pass these advantageous traits on, while traits that do not confer an advantage are not passed on to the next generation.
Go down a bit and the gap becomes even more glaring:
outcomes of evolution are sometimes divided into macroevolution, which is evolution that occurs at or above the level of species, such as extinction and speciation and microevolution, which is smaller evolutionary changes, such as adaptations, within a species or population.[141] In general, macroevolution is regarded as the outcome of long periods of microevolution.[142] Thus, the distinction between micro- and macroevolution is not a fundamental one – the difference is simply the time involved.
In short the origin of information challenge and the easily observed fact that multi-part function that depends on specific placement, connexion, fit and interaction of particular components is easily perturbed by removing or improperly adjusting key components, are being ducked. In short the general evidence points to islands of function, all across our world of experience. This extends to biology where sometimes as few as one or two changes in a protein can destroy function, as well as the factually observed reality of thousands of sharply distinct protein fold domains in the space of possible sequences, point strongly to islands of function. And, the proposed duck-out, of exaptation, runs smack into Angus Menuge’s challenges C1 – C5:
C1: Availability. Among the parts available for recruitment to form the flagellum, there would need to be ones capable of performing the highly specialized tasks of paddle, rotor, and motor, even though all of these items serve some other function or no function.
C2: Synchronization. The availability of these parts would have to be synchronized so that at some point, either individually or in combination, they are all available at the same time.
C3: Localization. The selected parts must all be made available at the same ‘construction site,’ perhaps not simultaneously but certainly at the time they are needed.
C4: Coordination. The parts must be coordinated in just the right way: even if all of the parts of a flagellum are available at the right time, it is clear that the majority of ways of assembling them will be non-functional or irrelevant.
C5: Interface compatibility. The parts must be mutually compatible, that is, ‘well-matched’ and capable of properly ‘interacting’: even if a paddle, rotor, and motor are put together in the right order, they also need to interface correctly.
( Agents Under Fire: Materialism and the Rationality of Science, pgs. 104-105 (Rowman & Littlefield, 2004). HT: ENV.)
Whether or not something is actually strictly irreducibly complex, these challenges have to be met for good engineering functionality reasons.
So, we can take islands of function to be a serious challenge, and a roadblock to the bland assertion that in effect macro-evo is nothing but accumulated micro-evo to the point where sufficient divergence of populations has occurred.
[U/d: Interview with Michael Behe on why he came to champion and still champions ID and irreducible complexity:]
[youtube reRTAD_6R84]
And, we have not touched on the observations made by S J Gould et al, on the problem posed by observed sudden appearances, stasis and disappearance, as a dominant feature of the fossil record at all levels. Of which, the Cambrian life revo is a capital case in point, where dozens of top level body plans appear suddenly in the record, in a window of 5 – 10 MY on the usual timeline. It would credibly take 10 – 100+ mn bits worth of functional info to do that, dozens of times over. Not to mention, this is TOP DOWN, not bottom up.
So, even if one accepts universal common descent — given the problem of cogently defining a species, every one including most modern YEC’s will accept limited common descent (probably up to more or less the family or equivalent level) — the issue of design as best explanation absent question-begging materialist a prioris has not been settled.
Perhaps the capstone case of reasoning in an enforced consensus-driven circle is this:
All organisms on Earth are descended from a common ancestor or ancestral gene pool. [–> begging the question] [173][245] Current species are a stage in the process of evolution, with their diversity the product of a long series of speciation and extinction events.[246] The common descent of organisms was first deduced from four simple facts about organisms: First, they have geographic distributions that cannot be explained by local adaptation. Second, the diversity of life is not a set of completely unique organisms, but organisms that share morphological similarities. Third, vestigial traits with no clear purpose resemble functional ancestral traits and finally, that organisms can be classified using these similarities into a hierarchy of nested groups – similar to a family tree.
Notice that implied “self-evident” point again?
Common design with built-in capability to adapt is at least as good an explanation of the facts in question as the underlying assumed naturalistic, blind watchmaker thesis evolution, but it has been excluded, a priori, and silently.
Within that circle, of course the only allowed explanation — and this is an inference to best current materialist explanation, not a deduction — degree of resemblance is suggestive of common ANCESTRY (after all, design has long since been ruled out), and the evidence of some adaptive radiation and biogeographic similarity — all of this well within the sorts of levels acceptable to even YEC’s — count as persuasive evidence of grand, cumulative divergence from the hypothetical common unicellular ancestor.
And, we still find no root to the tree that is empirically warranted.
So, common descent with divergence should be separated from the assumption that macro evo is simply cumulative micro, on the info-island of function challenge. That has to be bridged not dismissed. Next, common descent needs to be separated from universal common descent. And common descent to even universal degree in a world where common design is possible and compatible with such, needs to be separated from the assumption of blind watchmaker evolution.
Case not proved, and nowhere near being proved.
After 150 years, and counting.>>
_________________
All froth and no Maubee is the local saying (though I confess to not being fond of that bitter bark drink, much as I am not fond of my native country’s Blue Mountain Coffee).
Next, Theobald:
__________________
>> 6 –> There are several appeals to the true origins site, and to a series on 29 evidences of macroevo. The problems with Talk Origins as a site notorious for rhetorical manipulation are being ducked, and the essential problems with any macro evo blind watchmaker account absent demonstration of origin of FSCO/I by blind chance and mechanical necessity, are already outlined. In addition, we may look at the critique of the Theobald claims here, by Camp.
7 –> As one key point, observe the opening page of Theobald on his 29 evidences as linked: Universal common descent is a general descriptive theory concerning the genetic origins of living organisms (though not the ultimate origin of life).
Q: Why is the root of the whole tree so conveniently left off?
A: because there is no good empirical warrant for the claimed spontaneous origin of life in some chemical stew or other, and as a result apart from misleading icons, it is in a lot of trouble on the scientific merits. In addition, it is a capital illustration of the origin of FSCO/I, and of the failure to provide a viable blind chance and mechanical necessity mechanism. Where, ever since 1984, OOL has been the pivot of inferring design as credible best explanation of cell based life on the issue of origin of FSCO/I in light of empirically credible best explanation. Where also, once we see design as best explanation at this level, there is no good reason to lock out design onwards. That is, we here see a major begging of the question at stake in the teeth of knowing that this is the pivotal issue on the other side of the question. [Cf discussion of OOL at 101 level here on. Also, on OO body plans here on, including Gould’s remarks on the fossil record that is often alleged to show directly the universal common descent by BWE, here on in context, including of course the Cambrian revo as a capital illustration — never mind the dismissive remarks.]
8 –> As a second point, observe the following categorical blunder, on that same page:
universal common ancestry entails the transformation of one species into another and, consequently, macroevolutionary history and processes involving the origin of higher taxa. Because it is so well supported scientifically, common descent is often called the “fact of evolution” by biologists.
9 –> The main rhetorical lever here is that “only fools dispute facts,” and it is multiplied by a subtle no true scotsman [biologist] claim to collective authority.
10 –> Sorry, absent time travel, there is no possibility of actually observing directly the real remote past of life on earth. So, we are examining traces of the past and are seeking a “best explanation” in light of causal factors in the present observed to give similar results to a sufficient degree of closeness that we can identify empirically reliable signs and then infer per best explanation the credible causes. (Notice, how analogy gets embedded in inductive reasoning here. One of many ways this happens.)
11 –> Micro evo, as it is called, does not cross the FSCO/I body plan origin threshold, and is therefore not sufficiently parallel to properly conclude that the mechanism for the one is sufficient for the other. And, there is no observed case of origin of the required FSCO/I by accumulated micro evo etc, instead the only empirically known cause of FSCO/I is design.
12 –> So, what is happening is that big questions are being begged and on the false credit of institutional authority, so that something is being called a fact that is not. Patently, an inferred or claimed best explanation — even if FSCO/I had been warranted on empirical evidence, to come about by blind chance and mechanical necessity — would not be a fact in any proper sense. Scientific theories or models are explanatory constructs and on the history of science are inherently subject to overthrow in light of further evidence. Just ask the physicists on that. That is the category error involved.
13 –> So, this assertion (which is pivotal in Theobald) is all too clearly a blunder:
The worldwide scientific research community from over the past 150 years has discovered that no known hypothesis other than universal common descent [in context, on blind watchmaker mechanisms] can account scientifically for the unity, diversity, and patterns of terrestrial life. This hypothesis has been verified and corroborated so extensively that it is currently accepted as fact by the overwhelming majority of professional researchers in the biological and geological sciences
14 –> Instead, what is at the logically fallacious root of this false and unfounded perception of certainty has been highlighted by [Philip] Johnson:
For scientific materialists the materialism comes first; the science comes thereafter. [[Emphasis original] We might more accurately term them “materialists employing science.” And if materialism is true, then some materialistic theory of evolution has to be true simply as a matter of logical deduction, regardless of the evidence. That theory will necessarily be at least roughly like neo-Darwinism, in that it will have to involve some combination of random changes and law-like processes capable of producing complicated organisms that (in Dawkins’ words) “give the appearance of having been designed for a purpose.”
. . . . The debate about creation and evolution is not deadlocked . . . Biblical literalism is not the issue. The issue is whether materialism and rationality are the same thing. Darwinism is based on an a priori commitment to materialism, not on a philosophically neutral assessment of the evidence. Separate the philosophy from the science, and the proud tower collapses. [[Emphasis added.] [[The Unraveling of Scientific Materialism, First Things, 77 (Nov. 1997), pp. 22 – 25.]
. . . . PS: I again note that I have outlined how design can answer to the issues, here on at 101 level. In particular I there show why I confidently say that FSCO/I is an empirically reliable sign of design as credible cause.>>
I then went on to comment:
It should be clear as to why I am insisting that every tub must stand on its own bottom, and that claimed mechanisms of the past or explanations of the past must pass the test of causal adequacy relative to key traces, and further showing that the signs in question are able to distinguish candidates without begging questions . . . .
Why should I have any confidence in Theobald if in opening his discussion he cannot even get what is a fact straight? And, if those citing him so confidently cannot spot so gross a blunder, what does that tell us about the quality of their confidence?
A further full year later, I see no reason to take Theobald’s arguments seriously in light of so pivotal a blunder right at the outset. For those who want to wade through a point by point rebuttal, regardless of that stumble coming out the starting-gates, Camp’s reply here may be useful. It can be safely said that at no point does Theobald advance the case on the merits beyond the problems identified in the Wiki summary.
____________________
So, in the end, utterly unsurprisingly, it boils down to the obvious point that Lewontin’s a priori evolutionary materialism has become a dominant, deeply indoctrinated view:
. . . to put a correct view of the universe into people’s heads we must first get an incorrect view out . . . [–> worldviews level question-begging, leading to a Kulturkampf agenda of radical indoctrination in evolutionary materialism] the problem is to get them to reject irrational and supernatural explanations of the world [–> equating of accepting the reality of God to being irrational, an extremely arrogant and disrespectful attitude], the demons that exist only in their imaginations [–> demonising loaded language, and again question-begging], and to accept a social and intellectual apparatus, Science, as the only begetter of truth [–> NB: this is a knowledge claim about knowledge and its possible sources, i.e. it is a claim in philosophy not science; it is thus self-refuting] . . . .
It is not that the methods and institutions of science somehow compel us to accept a material explanation of the phenomenal world, but, on the contrary, that we are forced by our a priori adherence to material causes [–> another major begging of the question . . . ] to create an apparatus of investigation and a set of concepts that produce material explanations, no matter how counter-intuitive, no matter how mystifying to the uninitiated. Moreover, that materialism is absolute [–> i.e. here we see the fallacious, indoctrinated, ideological, closed mind . . . ], for we cannot allow a Divine Foot in the door. [–> exposure of inner spiritual motivation, fear and loathing of God leading to demonising him, viewing those who believe in him as irrational and “ignorant, stupid, insane or wicked” and more; and a declaration that science and science education are to be turned into propaganda front operations for an atheistical cultural agenda . . . no wonder atheistical activists are so desperate to suppress citation of this declaration of cultural war in the name of the science and science education elites.] [From: “Billions and Billions of Demons,” NYRB, January 9, 1997. Bold emphasis and notes added. If you have been led to imagine this is “quote mined” kindly cf the just linked fuller cite and notes. Also, in that context there are many more remarks that show this is not an isolated idiosyncratic view by an isolated individual, it reflects a major pattern and problem with both science and science education in our day.]
Philip Johnson’s reply to Lewontin is apt:
For scientific materialists the materialism comes first; the science comes thereafter. [[Emphasis original] We might more accurately term them “materialists employing science.” And if materialism is true, then some materialistic theory of evolution has to be true simply as a matter of logical deduction, regardless of the evidence. That theory will necessarily be at least roughly like neo-Darwinism, in that it will have to involve some combination of random changes and law-like processes capable of producing complicated organisms that (in Dawkins’ words) “give the appearance of having been designed for a purpose.”
. . . . The debate about creation and evolution is not deadlocked . . . Biblical literalism is not the issue. The issue is whether materialism and rationality are the same thing. Darwinism is based on an a priori commitment to materialism, not on a philosophically neutral assessment of the evidence. Separate the philosophy from the science, and the proud tower collapses. [[Emphasis added.] [[The Unraveling of Scientific Materialism, First Things, 77 (Nov. 1997), pp. 22 – 25.]
KS’ bomb has fizzled out. But in so doing, it has pointed back to the underlying problems with dominant origins science narratives presented to children in schools and the general public as fact, fact, FACT. Even, in part as self evident truths. It is time for a serious rethink and for fresh thinking.
As a final note, as there are threads where discussion can continue, e.g. here on where the grand body plan origin macroevolutionary and OOL narratives have come up, this will be FTR as a reference point.
There is plenty of room for discussion in and around UD, and the pro-darwinism essay challenge is still open [at two years, one month and counting . . . ], so claims of “censorship” and evasion of discussion or even cowardice on my or our part are patently groundless (Discipline for trollish misconduct is not censorship as there is no right to abusive comment.) If there was a dodge in this whole situation, it was the dodge on providing a simple 6,000 word essay supporting the evolutionary materialist paradigm, that could use any number of references and links elsewhere. And that dodge came from the design objector side, so this whole current programme of personalities can be seen as a turnabout accusation to distract attention from that embarrassing fact. Surprise — NOT. END